
1.Introduction
エビ・カニ類は甲殻綱(Class Decapoda)の中の十脚目(Order Decapoda)に属する。さらに,十脚目は大きく2つのグループ,つまり根鰓亜目(Dendrobranchiata)と抱卵亜目(Pleocyemata)に分類されている。根鰓亜目には,ウシエビやクルマエビなど比較的大型のエビ類が入る。一方,多くのエビ・カニ類は抱卵亜目に属する。根鰓亜目と抱卵亜目は,系統(phylogeny)をよく考慮しており,私はこの分類(classification)は妥当だと思う。
根鰓亜目のエビ類と抱卵亜目のエビ類については,外部形態(みかけ)はよく似ている。根鰓亜目のエビ類は,英語の通俗名はprawnになる。明確な定義がある訳ではないが,shrimpだと何か小さいエビ(小エビ)というイメージが付きまとう。根鰓亜目に属するshrimpがいるかは不明だが,現存していたとしても不思議ではない。なお,現存の小エビ類はすべて抱卵亜目に入る。
いわゆる「エビ」を区分する用語として,prawn(クルマエビを一般名称として使うことができる。),shrimp(小エビ),それにlobster(ロブスター)がある。分類学では,これらを十羽ひとからげで(「十羽ひとから揚げ」ではない。)エビ類と称している。エビという名称は,漢字で書ける(蝦,海老)が,なぜそんな古い名称が現代でも通用するのか,私にはよく理解できない。
ロブスターは,外敵に襲われたときには,後ろに泳いで逃げる。それ以外は,海底を歩行する。なお,「歩行する」の英訳はlocomoteやwalkになる。locomotionは「運動する・動く」というイメージが強い。人間臭い表現になるが,walkでいいと思うCrawl(這う)は良くない。「イザリガニ」のように卑屈な行動を連想させる。そんな名前の付け方はやめようではないか。
ロブスターは,一般名詞(prawn, shrimpに対するlobster)と固有名詞(Homarus americanusやH. norvegicus)の両方に使われる。日本語で言うならウミザリガニであるが,ザリガニはイメージが悪い。
海底や淡水の底を歩く十脚類(底生歩行エビ類)は,ザリガニ(crayfish)やイセエビ(spiny lobster)のように体の大きい種類もいるが,スナモグリやアナジャコのように小さい種類もいる。スナモグリはghost shrimp,アナジャコはmud shrimpと呼ぶ人が多いが,系統を考慮すると,両者はshrimp(小エビ)の仲間ではない。れっきとしたlobsterの仲間である。ロブスターというと外皮(exoskeleton)の硬い十脚類(イセエビとかゾウリエビとか)を想像するのだが,サンゴ塊の隙間,海岸に堆積した砂や泥に巣穴を作るようになってからは,外皮が薄くなる脱石灰化(decalcification)が進行した。脱石灰化は,十脚目甲殻類の進化を考えるうえで重要な指標になる。
系統を重視するならば,スナモグリはsand-dwelling lobster, アナジャコはmud-burrowing lobsterと呼ぶことができるだろう。周囲の同調を待っていたら,いつまでたっても我が道は開けない。ここは見切り発車で行かせていただく。
もう一度分類の話に戻ると,ロブスターと言われる十脚甲殻類は,いくつかの下目(infraorder)に分かれている。下目の名称にもしつこく「エビ」が付きまとっているので,できるだけ系統に近づけてみたい。アカザエビやザリガニは,原始歩行下目(Astacidea)。アメリカン・ロブスター(Homarus americanus)は原始歩行下目に属する。イセエビやゾウリエビは,岩礁歩行下目(Palinura)に属する。アナエビ類(Axiidean lobsters)もロブスターの一種であり,内生型歩行下目(Axiidea)という名称をつけてみた。異尾下目(Anomura)および短尾下目(Brachyura)は,そのままでよいか・・・。
内生型歩行類(下目)に属するアナエビ類(Axiidea)は,いくつの目(order)で構成されているのか不明である。最近の分類では,アナエビ下目にスナモグリが入ってきている。しかし,こんな分類ではとても収拾がつかなくなる。分類学(taxonomy)では,分類の基準(standard)が,客観的な指標(例えば分子系統解析や特定の構造や器官の有無)に準拠するのではなく,個人(自称権威者)のさじ加減によって決まる。権威者が変われば,それぞれの生物の分類学的な位置づけ(taxonomic position)もコロコロと変わる。そんな学問もあるかもしれないが,私は従いたくはない。
アナエビ類(Axiidean lobsters)がいくつの目(order)や科(family)から成るのか,現状では定まっていない。アナエビ類全体を見渡すと,みんなよく似ている。もっと正確に言えば,それぞれのグループを分ける具体的な指標が見当たらない。分類学者はあるというだろうが,思いこみが先に立ってエビデンス(evidence)がともなっていないから,客観性にも乏しい。
そもそも,内生型歩行類(下目)自体が,原始歩行類(ザリガニ)や異尾類(ヤドカリ)と同列の分類学的地位(status)にあるのか?それも不明である。アナエビ類というのは,他の下目の十脚甲殻類に比べて,映え出た形態的特徴がないのが特徴と言える。なぜ際立った特徴がないのかというと,それはアナエビ類の進化が深く関係しているように思われる。
アナエビ類は,アカザエビ(Metanephrops)やロブスター(Homarus)の祖先のような原始歩行類から直接進化したのだろう。例えば,原生のロブスターが何年生きるかわからないが,多くの文献を見ると,最低でも10年は生きそうだ。20年から30年なんて個体も普通にいる感じがする。
野生の生物は,環境条件が良ければ,私たちが想像しているよりずっと長く生きることがわかっている。アカテガニは10年以上生きた記録がある。ブッポウソウは,10年以上は確実に生きる。20年生きるかもしれない。ぴよ吉がそれを実証してくれるかもしれない。
一方,アカザエビやロブスターが長生きするからと言って,成熟するまでの年数も長いとは限らない。抱卵亜目の十脚類では,排卵(ovulation)と同時に受精が起き,受精卵は腹部の体表に生えている担卵毛(ovigerous seta)に付着する。それぞれの卵(受精卵)では胚発生が進行し,幼生(ゾエア幼生)の形態ができあがるとふ化(hatching)が起きる。
ふ化は,受精時の卵の表面から分離,肥厚した卵殻が破裂することで起きる。ゾエア幼生は,6回の脱皮を経て,バカでっかい鋏脚を持ったロブスターの形(young)に少しずつ近くなってゆく。以降,脱皮回数は年数とともに減少する。雌雄とも性成熟が起きるのは,ふ化してから4~5年はかかると予想される。
話は脱線する。ロブスターは腹部にもぷりぷりとした身があるし,鋏脚,特に右側の鋏脚は巨大で,中にはおいしい身がたくさん詰まっている。十脚甲殻類の中でも,ロブスターの筋肉の肉質と体全体に詰まっている肉の量は,トップクラスに近い。ただし,ロブスターの形状はザリガニと似ている。確かにザリガニはロブスターの祖先から進化したには違いないが,両者の系統的位置は大きく離れている。ザリガニの仲間というイメージで食べないことが大事かもしれない。
ロブスターの胚発生や成長に関しては,100年以上も前から研究が行われている。ロブスターの成長(growth)に関する文献を見ていると,面白いことに気付かされる。ロブスターの特徴は,何と言ってもあの大きな左右の鋏脚である。一方,成長段階にある若いロブスターはどうか? ロブスター固有のバカでっかい鋏(はさみ)はまだなく,鋏脚(かんきゃく)の先にはザリガニよりもまだ小さい鋏がついている。そのため,個体全体を眺めると,ロブスターではなく,アナエビの一種のように見える。
ロブスターが成長して行く過程で,ふつうは4~5年で性成熟が起きる。しかし,サンゴ塊の間に多く隠れ場所のある(隙間のある)場所に移り住んだ個体の中に,まだ若いうちに(例えば,ふ化後2年目)に性成熟が起きる突然変異が生じた可能性がある。早熟を引き起こす突然変異遺伝子は,グループ全体に広がり,アナエビ(原始アナエビ)が誕生したのだろう。こうした現象は昔からネオテニー(幼形成熟)として知られていて,生物進化のひとつのパターンである。
アナエビ類の分類に戻る。十脚甲殻類の他の分類群を考慮するならば,アナエビ類を内生型歩行下目(目と科の中間)に置くことは可能である。アナエビの祖先は,多くは絶滅しただろうが,現生のアナエビはサンゴ礁原に多い。現状では形として2~3の目が設定できるだろう。ヤハズアナエビのようにサンゴ礁原の石灰岩に穴を穿つ礁原穿孔科,ミツトゲイソアナエビのようにサンゴ塊の隙間にすむサンゴ塊穿孔科,それにシロオビアカアナエビやソーヨウアナエビのように,泥干潟や海底に穴を掘る砂泥底穿孔科ぐらいを設定したらどうだろうか。それぞれの科(family)の中に自分が思った属(genus)を散りばめれば,アナエビ類の分類は完成する。
誰とは言わないが(言ってもいいけれど・・・),日本人,外国人とも,権威が入り込むと,生物の分類はおかしくなる。(おかしいのはお前の方だろう,というご意見も多いかと思うが,ここは私の立場で書いている。)権威の方々にはここはぐっと我慢し,ご遠慮いただくようお願いしたい。ここから先は,権威の方々のご教示(多くはお怒りの言葉)がなくても戦える。権威者の方々には大所・高所から,新規参入者(新規侵入者?)を温かく見守っていただけるとありがたい。
アナエビ類を固有の下目に置かないで,他の下目のもとに置くと大きなトラブルに発展する。トラブルが起きると権威者が降臨してせっかく立ち上がった若い芽もすぐに摘み取られる。アナエビ類を固有の下目(内生型歩行下目)に置くのは譲れない。
私たちの分類基準では,分子系統学的知見が重視される。これからパブリッシュする論文では,十脚甲殻類の分類と進化を論じる際には,必ず分子系統解析の結果を提示して意見を述べることとしたい。
アナエビ類は,日本列島沿岸だと,相模湾の深海に1種類(ソーヨーアナエビ)分布する。ソーヨーアナエビは,沖合底引き網で採集されるから,アカザエビのように普段は海底に作られた巣穴にいて,ときどき巣穴を抜け出して海底を歩き回る生活を送っているのだろう。
日本列島沿岸の深海にはまだ数種類か生息していると思われる。瀬戸内海には多分アナエビ類はいない。日本海側には,ソーヨーアナエビは分布していると思う。土佐湾では,司丸(19 t)のご協力を得て,水深150m~350mの海底でたくさんの十脚甲殻類を採集した。また,第17宝生丸(4.9 t)では深さ30m~60mの海底で十脚甲殻類を採集した。いまのところ,どちらの採集でもアナエビ類は土佐湾の深海からは獲れていない。
アナエビ類(内生型歩行下目)が多いのは,琉球列島である。琉球列島には広大なサンゴ礁原が広がり,サンゴ礁原や礁原上の(死んだ)サンゴ塊の隙間にいろいろな種類のアナエビが生息している。サンゴ礁原は場所によっては泥干潟になっている。ベイトポンプを使って丹念に泥干潟を調べると,数は少ないが,アナエビを採集することができる。泥の中に棲んでいるのに,外皮は大変カラフルである。
シロオビアカアナエビとシロオビアオアナエビについてはすでに紹介した。本記事においては,ヤハズアナエビを紹介したい。なお,亜熱帯域では底引き網漁はやっていないので,深海にはどんなアナエビが生息しているか不明である。・・・でも,多分少ないと思われる。
2.撮影と執筆の基本情報
<撮影場所> 西表島と石垣島(採集:三枝誠行,南里敬弘)。<撮影と記事執筆> 三枝誠行(生物多様性研究・教育プロジェクト常任理事)。
<Key words> 内生型歩行下目,Axiidea,アナエビ類,ヤハズアナエビ,琉球列島,サンゴ礁原,ロブスター,礁原穿孔科。
3.参考文献
・Castro, P., and M.E. Huber. 2005. Marine Biology (fifth edition). McGrow Hill Higher Education.
・Factor, J.R. ed. 1995. Biology of the Lobster Homarus amaricanus. Academic Press.
・Futuyma, D.J. 1998. Evolutionary Biology. Third Edition. Sinauer.
・Herrick, F.H. (1895) The American lobster. A study of its habits and development. Bull. United States Fish Comm. 1895: 1-252 with 16 plates.
・Kensley, B. (1989) New genera in the thalassinidean families Calocalididae and Axiidae (Crustacea: Decapoda). Proc. Biol. Soc. Wash. 102: 960-967.
・Kensley, B., and Williams, A.B. (1990) Axiopsis eximia, a New Thalassinidean Shrimp (Crustacea, Decapoda, Axiidae) from the Middle
Eocene of South Carolina. J. Paleontol. 64: 798-802.
・Kensley, B. (1996) New thalassinidean shrimp from the Pacific Ocean (Crustacea: Decapoda: Axiidae and Calocaridae). Bull. Mar. Sci. 59: 469-489.
・Kensley, B., F.-J. Lin, and H.-P. Yu (2000) Further records of thalassinidean shrimps from Taiwan (Decapoda: Axiidae and Calocarididae), with descriptions of three new species. J. Crust. Biol. 20 (2): 207-217.
・木村資生 1988. 生物進化を考える。岩波新書。
・Saigusa, M. (1992) Observations on egg hatching in the estuarine crab Sesarma haematocheir. Pacific Science 46: 484-494.
・竹山佳奈・他7名(2014)西表島北西部における熱帯性海草ウミショウブの広域分布および生育条件に付いて。土木学会論文集B3(海洋開発)70: 1068-1073.
・Wikipedia:ネオテニー(https://ja.wikipedia.org/wiki/ネオテニー)
4.アナエビ下目(穿孔歩行下目)の分類(Kensley and Williams 1990)
Order Decapoda, Latreille 1803 ・・・・・・ 十脚目
Suborder Pleocyemata, Burkenroad 1963 ・・・抱卵亜目
Infraorder Thalassinidea, Latreille 1831・・・・タラシナ下目(穿孔下目)
Superfamily Thalassinidea, Latreille 1831・・・タラシナ上科(穿孔上科)
Family Axiidae, Haxley 1879・・・・・・・・アナエビ科(穿孔科)
Genus Axiopsis, Borradaile 1903 ・・・・・・砂泥底穿孔属
Species Axopsis eximia, Kensley and Williams 1990(種名)
5.アナエビ類(内生型歩行下目)の分類(Saigusa 2023)
Order Decapoda, Latreille 1803 ・・・・・・ 十脚目
Suborder Pleocyemata, Burkenroad 1963 ・・・抱卵亜目
Infraorder Axiidea・・・・・・アナエビ下目(内生型歩行下目)
Superfamily Axiidea ・・・・・アナエビ上科(内生型歩行上科)
Family Axiidae, Haxley 1879・・アナエビ科(礁原穿孔科)
Genus Neaxius ・・・・・・・ 砂泥底穿孔属
Species Neaxius acanthus・・・・ヤハズアナエビ(種名)
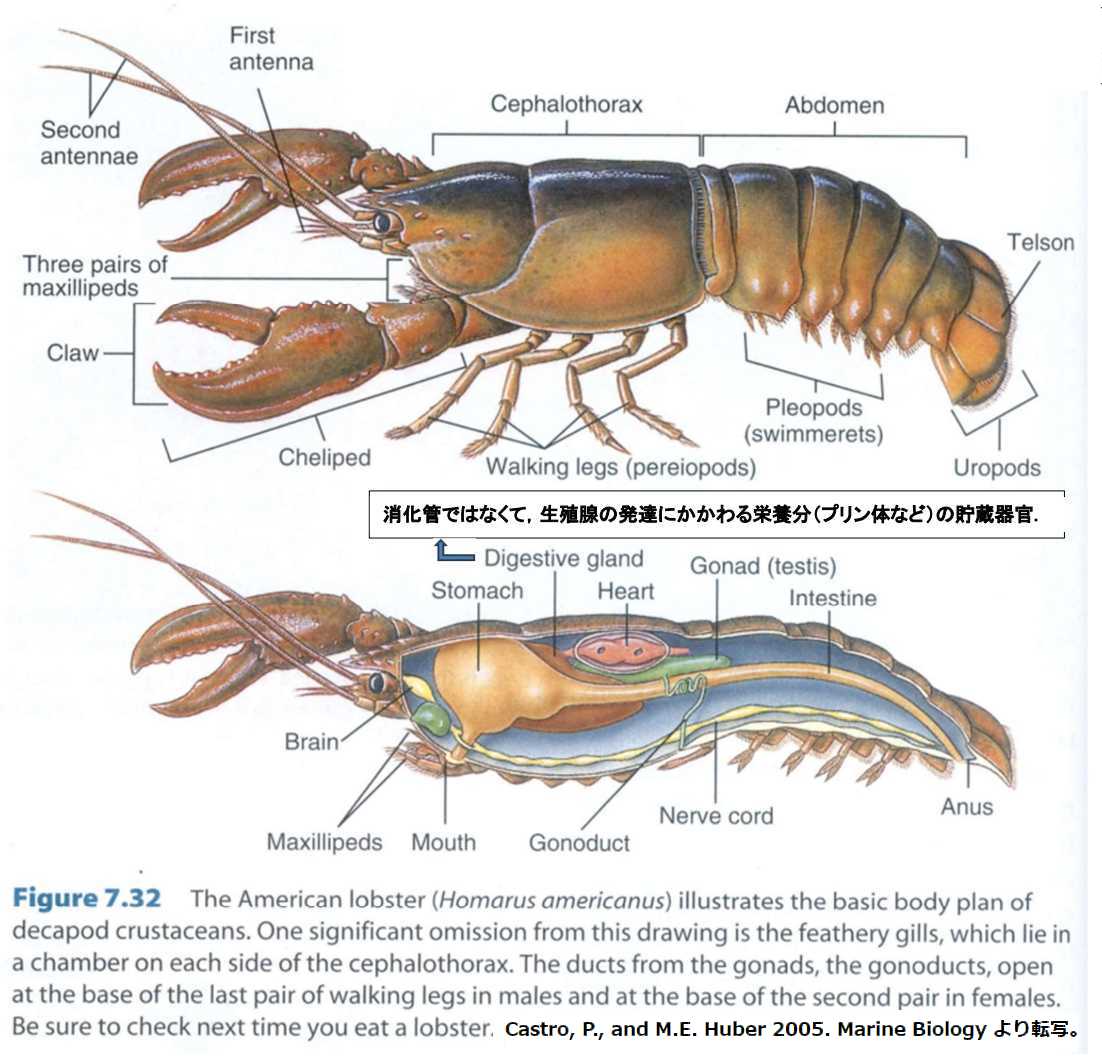
図1.ロブスター(ウミザリガニ)の外骨格構造。(上)外骨格。(下)内部器官。内部器官は血管系、神経系、消化管、生殖腺。肝臓と膵臓はない。

図 2.アメリカン・ロブスター。左がオス個体で,右がメス個体。左右どちらの鋏脚(かんきゃく)も,メスよりもオスが大きい。Herrick, 1895 の論文から転写。この写真は,1890 年前後に臨海実験所(Woods Hole?)で撮影されたのだろう。そのころの日本では,1877 年創立の東京大学が,1886 年帝国大学に改称された。同じ年に,三崎臨海実験所が設立された。破門草事件(伊藤篤太郎が,矢田部良吉に内緒でトガクシソウの新しい学名(新属)を発表し,帝国大学植物学教室への出入りを禁じられた事件)が起きたのもこの頃である。さらに(私とは絶対馬が合わない)丘浅次郎が帝国大学理科大学動物学科選科に入学したのは,1886 年である。一方,アメリカでは,1890 年ごろにはこんな素晴らしい写真が撮影できるほど,科学は進歩していた。
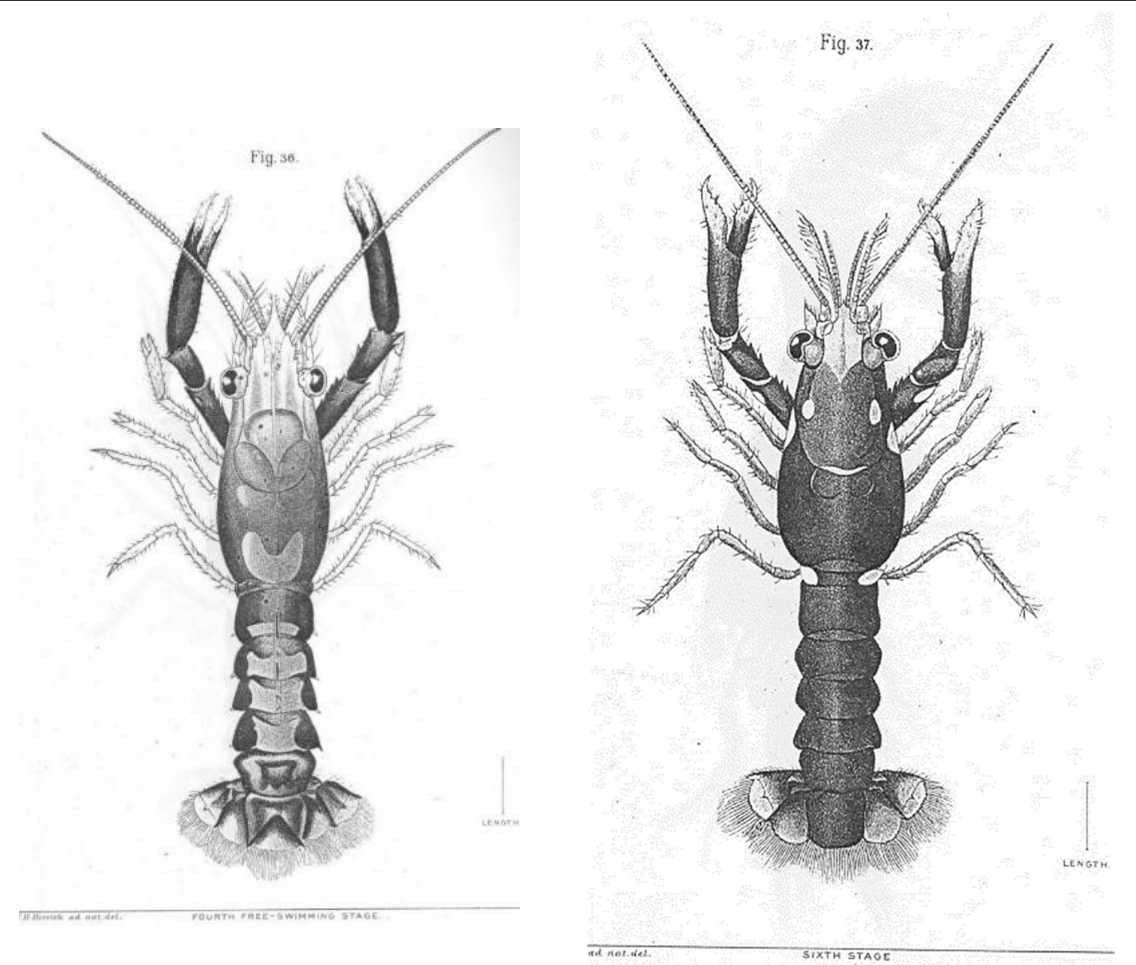
図 3.(左)アメリカン・ロブスターのふ化後 4 令(4 回脱皮したという意味)の若い個体。(右)ふ化後 6 令の個体。アメリカン・ロブスターの胚発生期間は 1 か月ぐらいあるだろうか。ふ化(卵殻が破裂して幼体が外に出る)後は,3 期の幼生を経て 4 令の幼体になる。
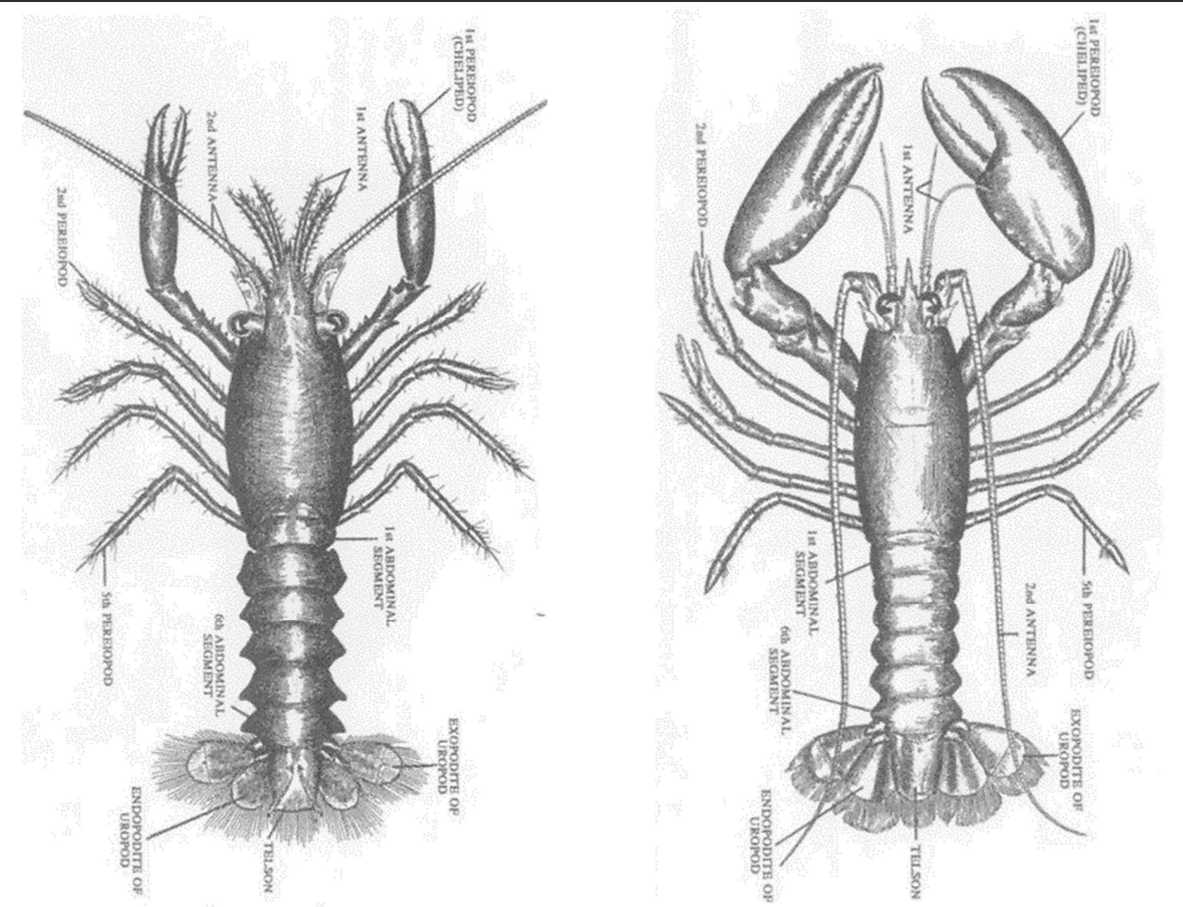
図 4.(左)アメリカン・ロブスターのステージ 4 期(4 令)の幼個体。図 3 に示した 4 令の個体と同じ成長のステージ。(右)同ステージ 14 期(14令)の若い個体。Factor, J.R. (1995) Introduction, anatomy, and life history. In: Chapter 1, ‘Biology of the Lobster Homarus americanus’. Academic Press. Figure 7 を転写。こちら(図 4)はスケッチ。ロブスターの 4 令期の幼体(young)の外形は,琉球列島で採集できるアナエビ類(Infraorder Axiidea)によく似ているように思えるが,どうだろうか?ロブスター(Homarus)は,アナエビ類の直接の祖先であるように思われる。

図 5.ロブスターの祖先と並んで,アナエビ類(Axiidea)の進化の母体となった原始歩行下目の十脚類がいる。それは,アカザエビ(Metanephrops japonicus)の祖先(ancestor)である。多くのアナエビ類の頭胸部の背面には,鋭い棘(とげ)の名残がある。アカザエビの祖先も起源の可能性がある。
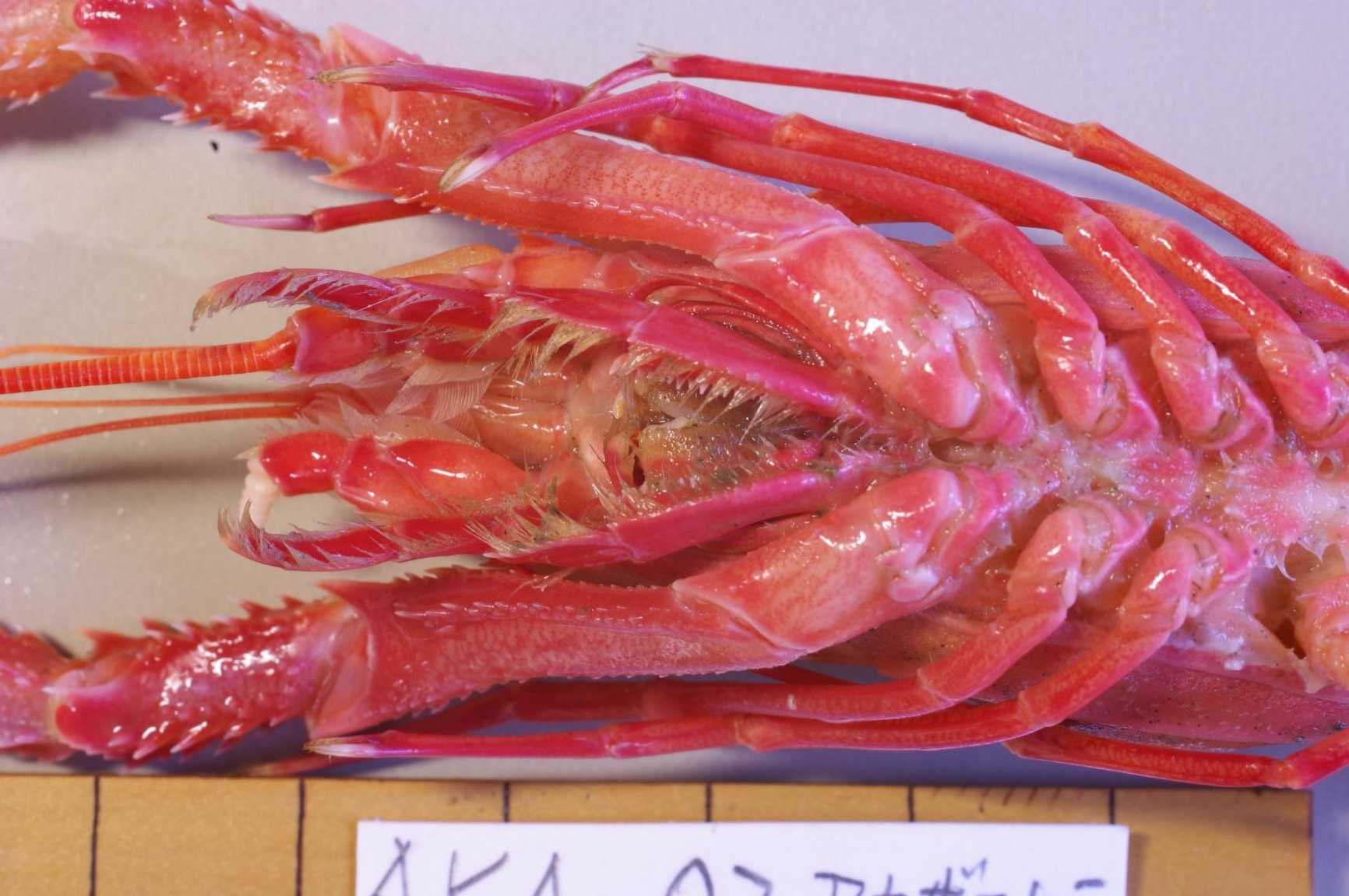
図 6.アカザエビの胸部腹面。第 3 顎脚の付け根の後方,鋏脚の付け根の前方には隔壁はなさそうである。・・・が,もう少しよく調べてみたい。

図 7.アカザエビの胸部腹面。アカザエビの面白いところは,鋏脚の付け根(基節)は接しているが,第 2 歩脚より後は離れていることである。
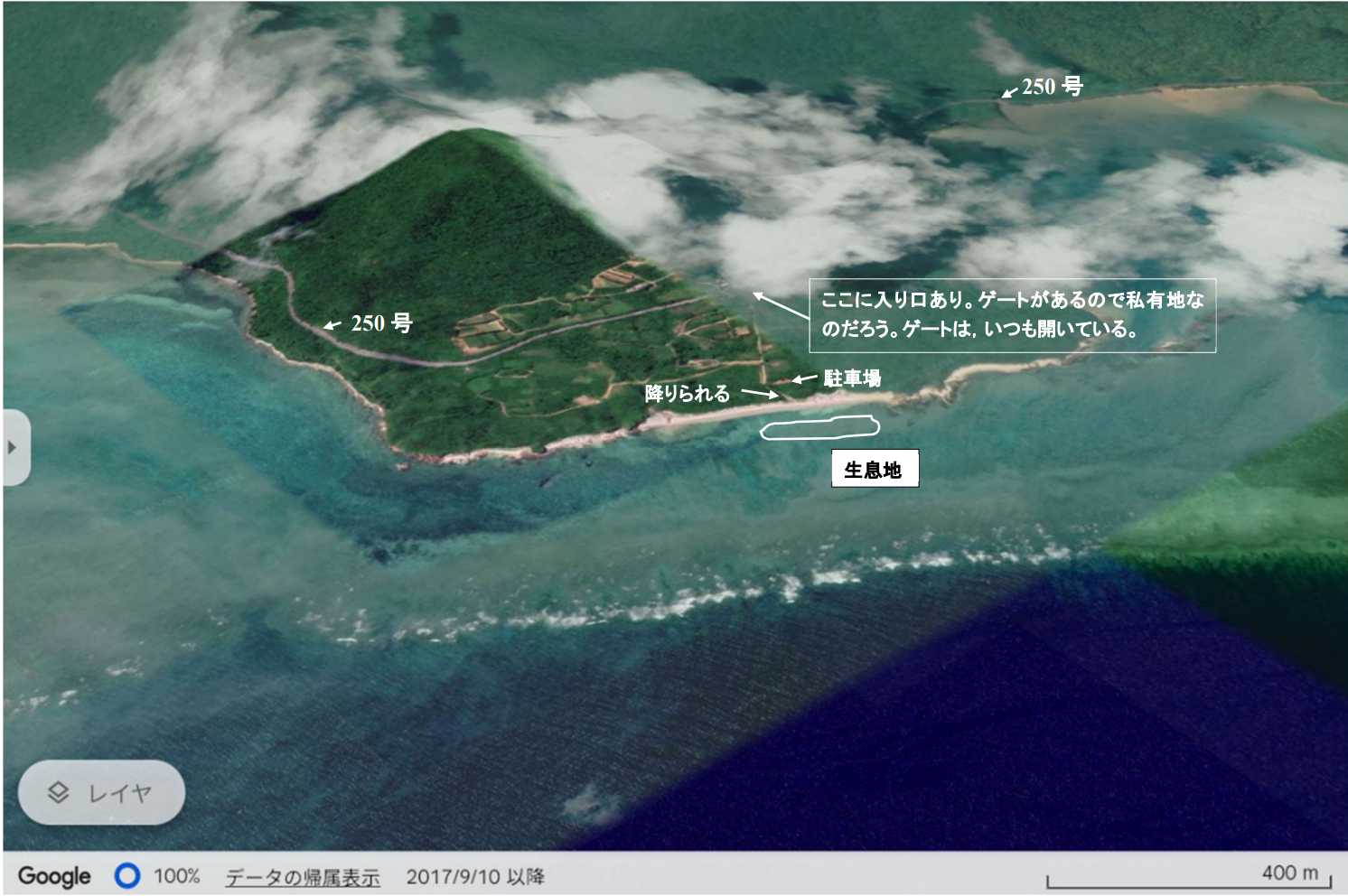
図 8.ヤハズアナエビの生息場所(赤離の海岸)。県道 250 号の道路わきにあるゲート(いつも開いている)から入る。100m 程のところに駐車場あり。

図 9.ヤハズアナエビの生息場所。波打ち際から 20-30m のところに浅瀬(ほぼ大気中に露出)がある。ヤハズアナエビはサンゴ礁原に穴を穿っている。

図 10.ヤハズアナエビ。巣穴は,サンゴ礫の隙間に作られているので,穴の周囲は死んだサンゴ塊なので,いったん穴に引っ込んだら採集は難しい。

図 11.サンゴ礫の隙間に作られた巣穴の中にいるヤハズアナエビ。鋏の力は弱い。ヤハズアナエビは第 2 歩脚もハサミ脚になっている。

図 12.ヤハズアナエビの頭胸部(背面)。額角(rostrum)と「つぶれ U 字溝」の間は平坦で棘(とげ)がない。ロブスターの頭胸部(背面)とよく似ている。ヤハズアナエビの祖先は,アカザエビよりもむしろロブスターの祖先と系統的に近いと予想される。

図 13.ヤハズアナエビの頭胸部(背面)。額脚(rostrum)と「つぶれ U 字溝」の間には棘がない。ヤハズアナエビの学名は Neaxius acanthus。礁原穿孔科のアナエビ(Neaxius)は種類が非常に少ないと思う。ヤハズアナエビの祖先は,原始歩行下目のロブスター(Homarus)の祖先と近いのではないか?

図 14.大潮の干潮時のリーフの縁(赤離:アカパナリ)。波は穏やかだが,サンゴ礁の縁(edge)は複雑な構造になっているので,歩くときには十分に注意。このあたりだと生きているサンゴや褐藻類は無数にある。小学生以下の子供は連れてきてはダメ。遠くに見えるのは 1960 年ごろに難破した貨物船。

図 15.サンゴヒメエビの一種。鋏脚は第 1 歩脚ではなく,第 3 歩脚であることに注意。第 2 歩脚の先端もハサミ状になっているようだ。第 1 歩脚と第 2 歩脚は顎脚(第 1 から第 3 顎脚)と同じような機能を持っていると予想される。なぜこのグループだけ第 3 歩脚が大きな鋏脚になったのか,不明。

図 16.アカパナリの海岸では,貝類やタコがたくさん捕れる。干潮時を過ぎて潮が上げ始めるころに,獲物を持って引き上げてくるおばちゃん。獲物はイモガイ(Conus)が多いと思う。おばちゃんたちは,リーフの縁からどこを通れば岸に戻れるかよく知っている。私はよく知らないので深みにはまって難儀することが多い。写真右上の船は西表島と石垣港を結ぶ高速船(上原航路)。石垣港に向かっている。波は穏やか。2016 年 4 月 9 日。

図 17.イボバサミアナエビ。鋏脚にイボイボのあるアナエビ。アルコール標本になってしまったが,生きているときにはきれいな色をしていた。どこかで写真を見た記憶があるが,思い出せない。属は Calocarides ではなく,Ambiaxius なのだろうか?アナエビの属や科はよくわからないが,分類学者に尋ねると大きなトラブルは避けられない。過去に多くのトラブルがあった。自分で判断するしかない。西表島干立の前の浜で採集した。個体数は非常に少ない。

図 18.シロオビアカアナエビ。これは Axiopsis serratofrons で間違いない。シロオビアオアナエビは,リーフの縁に近いサンゴ礫の底質に巣穴を作るが,シロオビアカアナエビの方は,外洋に面するといっても,やや内湾的な海岸にいる。つまり,陸上のすぐ近くで(10m ぐらいのところ)で採集される。

図 19.私が見ている形態学的特徴は分類学者のそれとは大きく異なる。たとえば,2 種類のアナエビ類を比較して,分類学者は違いを見つけるが,自称進化生物学者である私は,両者の共通した特徴を見る。日本の社会は,分類のようなことに興味を持っている人たちが多い。ブッポウソウでは,きれいな野鳥を自分も見てみたい,撮影してみたいという人は多いが,ブッポウソウという鳥がどんな習性を持っているのかについて関心のある人は少ない。同様に十脚甲殻類についても,食べることとか分類のこととかに興味を持つ人は多いが,生態・行動・進化について真面目に考える人は少ない。

図 20.シロオビアカアナエビの頭胸部(背面)。小さな棘が中央に 1 列,外側に左右 1 列,その内側に左右 1 列に並ぶ。これはサンゴ姫エビにも共通する特徴である。このような割と多くの十脚甲殻類に共通する特徴は,原始歩行下目のアカザエビの祖先から由来したと考えることはできないだろうか。

図 21.ハサミシャコエビ。琉球列島から本州の河口(estuary)の泥干潟(mud-tidal flat)には,泥を盛り上げて「塚」を作り,塚に穴を開けてその中に住んでいる十脚甲殻類がいる。ハサミシャコエビもまたロブスターの一種である。ハサミシャコエビ,オキナワアナジャコ,イシアナジャコ,ドロアナジャコ,スナモグリなどは海岸や河口域の砂泥底に巣穴を作って住む十脚類であるが,分類が定まっていない。従来は Thalassinidea にひとまとめにしてあったが,原始歩行下目の次に古い内生型歩行下目(アナエビ類)は,Thalassinidea から独立させた。アナエビ類に起源をもつ後発グループをどうまとめるか?

図 22.ハサミシャコエビ。採集してからすぐに撮影したので,泥がいっぱいくっついている。外皮(exoskeleton)は,イシアナジャコ類,アナジャコ類,スナモグリ類に比べて石灰化が進んでいないので,体表はやや硬い。ロブスターの雰囲気が残る。いままで私は,イシアナジャコ類,アナジャコ類,スナモグリ類と並んでハサミシャコエビがどこから来たのか理解していなかったが,アナエビに関心を持ってから,ようやくわかるようになってきた。

図 23.オサテエビ(原始歩行下目)。ハサミシャコエビとオサテエビの形態を比較すると,互いに非常によく似ていることが分かる。だからと言って,ハサミシャコエビが,オサテエビの祖先から進化したとは言えない。オサテエビの鋏脚は,左右で異常に特化した形態になっている。一方,ハサミシャコエビの鋏脚は多くの十脚甲殻類の鋏脚の構造に近い。アカザエビの祖先(原始歩行下目)の 1 種が,サンゴ礁に進出し,三畳紀末には砂泥穿孔類として礁原の砂泥底に生活していたのだろう。ジュラ紀と白亜紀は,形態とハビタットは,ジュラ紀と同じだったが,新生代になり富栄養化の進んだ河口域に進出したのだろう。分類としては,私はハサミシャコエビやオキナワアナジャコは Thalassinidea(Neothalassinidea はどうか?)に入れてよいと思う。

図 24.ハサミシャコエビ(Laomedia astacina)? 石垣島名蔵のマングローブで採集(ベイトポンプ)。2005 年 5 月 27 日(採集者:南里敬弘)。私は長いこと,この個体はアナエビの 1 種と思っていたが,ハサミシャコエビの小さい個体かも知れないと思い始めた。しかし,まだアナエビの可能性は残る。分子系統解析をすれば明確になる。どっちに転んでもハサミシャコエビは,アナエビ類(内生型歩行下目)に系統的に近縁だろうとは言える。

図 25.スナモグリの一種(種名不詳)。スナモグリの仲間は,サンゴ礁の砂泥底から海岸や河口の砂泥底に移る際に,外皮の脱石灰化(decalcification)にともない,形態の変化も進んだ。U 字型の溝から頭部先端にあった多くの棘や,棘群の名残も完全に消失したのだろう。にもかかわらず,体の基本的な構造はロブスターであるアナエビ類(内生型歩行下目)の特徴をよく残している。2005 年 5 月 27 日,石垣島名蔵湾で採集した(南里敬弘)と思う。

図 26.スナモグリの一種(種名は不詳)。古生代末に起きた海産無脊椎動物の大量絶滅(95%以上が絶滅)の後,中生代の三畳紀に徐々に生物相が回復した。生物相回復の基盤になったのは,古生代とは違ったタイプのサンゴ礁(coral reef)だったように思える。テチス海に始まったサンゴ礁は,大陸の分裂とともに地球上に広く分布を広げた。サンゴ礁の分布拡大に呼応して,サンゴ礁と周辺の環境で生活を共にした十脚甲殻類も,属,科,目のレベルで多様化してきたと考えることができる。中生代から新生代にかけてのサンゴ礁と十脚甲殻類の共進化(coevolution)は地球環境を大きく変化させただろう。