
2024 年 1 月 3 日(水)
1.Introduction
イシアナジャコは,西表島の美田良で初めて発見されてから,海岸や河口の波打ち際に転がっている砂泥岩(sandy-mud stone)の中に高密度で棲んでいることが分かった。なお,砂泥岩は造語であり,巣穴のある石や岩の粒度分析をすると,砂岩と泥岩のちょうど中間の粒度組成であることから名づけられた。砂泥岩はわりに柔らかく,ノミとトンカチがあれば簡単に崩すことができる。
イシアナジャコは,西表島では島の西側から北側の海岸にしか分布しない。例外として,東側の後良川を横切る橋(後良橋)の真下にある砂泥岩にも棲んでいるが,これは橋を作るときに橋げたを固めるために人為的に置いたものだ。
イシアナジャコが,なぜ西表島の西側海岸にしか分布しないかには,明確な理由がある。イシアナジャコは,海岸の波打ち際に転がっている砂泥岩にのみ生息している。アナジャコの棲める砂泥岩は,島の西側から北側にかけての海岸沿いに分布しているからである。
海岸の崖から転がり落ちるか,露頭として海岸に押し出された砂泥岩は,波に洗われて,細かい粒子に分解するか,あるいは海岸の砂浜に潜って行き,20 年もすれば消滅する。イシアナジャコが巣穴を掘ることのできる砂泥岩は,常に波打ち際に供給される必要がある。
島の東側の海岸は,隆起したサンゴ塊(死骸)か,非常に硬い砂岩(sand stone)によって占められている。波打ち際ではサンゴ塊や硬い砂岩の隙間に入り込んでいる生物はいるが,巣穴を作ることのできる生物はいない。・・・ということで,イシアナジャコは,島の東側の海岸には分布せず,西側から北側半分の潮間帯に限られている。
古くから分類学やってきた人たちは,生物を分類するのに直感(intuition)を頼りにしている。リンネ(Carl von Linné; 1707‒1778)の提唱した方法をそのまま引き継いでいる。一方,最近発達した分子系統解析によると,形態のスケッチでは区別できない種や属があることがわかってきた。形態的に区別できない種や属を,分子系統解析の結果だと言って,分類学者に新しい名前を付けろとは言えない。しかし,分類学者の判断を待っていては,私たちの研究が止まる。
2.撮影と執筆の基本情報
<写真の撮影と記事の執筆> 三枝誠行(生物多様性研究・教育プロジェクト常任理事)。
3.参考文献
・Castro P., and M.E. Huber (2005) Marine Biology, Fifth Edition. McGraw Hill Higher Education.
・Futuyma, D.J. (1998) Evolutionary Biology, Third Edition. Sinauer Associates, Massachusetts.
・Nibakken, J.W. (2001) Marine Biology, An Ecological Approach, Fifth Edition. Bemjamin Cummings, San Francisco.
・Ferl, R.J., and R.A. Wallace (1996) Biology: The Realm of Life, Third Edition. Harper Collins College Publishers.

図 1.西表島の西海岸。矢印はイシアナジャコ類が棲む海岸。イシアナジャコは入り江や河口の波打ち際にある砂泥岩に巣穴を作っている。

図 2.美田良(矢印 2)の浜。もう 20 年前になるが,私が初めてイシアナジャコを採集した海岸。台形の中にある砂泥岩をトンカチで割ったときに出てきた。その後,イシアナジャコは西表島の西海岸に多数生息していることがわかり,2023 年現在で西表島では 5 種類が新種として記載されている。
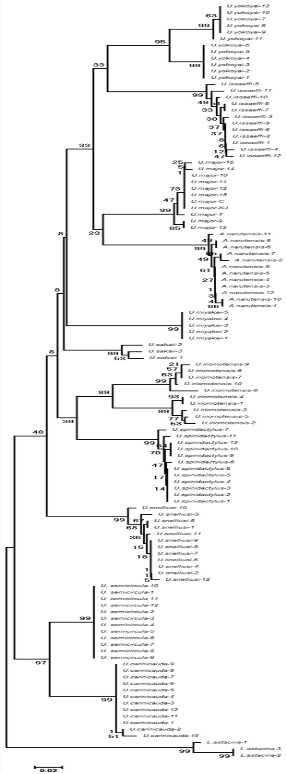
図 3.日本列島の沿岸の干潟に棲む 14 種類のアナジャコ類(Gebiidae)の分子系統解析。
種名は文字(letter)が小さすぎて私も判読できないが,細かいところは無視してよい。大事なのは,いままでアナジャコ類は全部 Upogebia 属に入るとされてきたが,分子系統解析の結果からは,外群を除いて 7‒8 属が区別されることである。それでは,誰が新しい属に対して名前を付けるのかという大問題が生じる。直感で生物の種類を判断する分類学者は,間違いなく逃げてしまう。アナジャコのように,底質の中に巣穴を作って棲む生物では,形態の収れん(convergence)が激しく,直感では属を分けられないからである。直感で判断できないのだから,当然名前もつけようがない。
A 属,B 属,C 属・・・と記号をつけることは可能である。あるいは,無理に固有名詞をつける必要はないという考え方もある。しかし,新属の可能性が高いのであれば,ABC・・・の代わりに生息場所の環境に関わる名前(例えば Stonogebia)をつけた方が,論文を見る方には理解されやすい。
なお,ナルトアナジャコ(和名)だけは Austinobebia という別属にされていた。形態から見ても,本種だけが別属に置かれるのはどう見てもおかしい。実際に分子系統解析を行ってみると,別属という判断は全く支持されないことがわかる。遺伝子解析の結果では,本種(Austinobebia)は上から 5 段目の種のグループ(図 3)に入っている。少し見にくいが,「A. narutensis」というグループがあるのがわかるだろう。分子系統解析の結果では,A. narutensis は U. major の隣のグループに入っていることがわかる。U. narutensis と U. major を合わせて,例えば Macrogebia 属とすることが可能だろう。
なお,図 3 ではどの属も種名までは判別できないが,属名や種名をわざとぼかしている訳ではない。図自体が縦長で大きく,縮小すると細かい部分まで見えなくなってしまう。もっと見やすいように,早急に図 3 を改善する必要がある。
ナルトアナジャコは,形態学的には容易に区別できる。ナルトアナジャコには第 6 腹節(尾節)の付属肢外肢の縁に小さな棘がある。初めて見る人は瀬戸内海の泥干潟にいるアナジャコ(Upogebia major)と区別がつかないが,慣れるとわかる。私は両者の区別は,生息場所の底質(substrate)で行っている。アナジャコはぬたぬたした泥に穴を掘るが,ナルトアナジャコは砂地の泥に穴を掘っている。瀬戸内海では,本州側の海岸には粒度の細かい泥(いわゆるヘドロ)が堆積するが,四国の海岸は砂泥底の浜が続く。ナルトアナジャコは砂泥底(四国側)の浜に生息する。ナルトアナジャコの場合には,命名者の分類に対する基本的な知識が著しく欠如した結果と言わざるを得ない。
遺伝子解析の結果は,アナジャコとナルトアナジャコのどちらも,種としては良くまとまっている。命名者は,属が違うほど類縁関係が離れていると思ったのだろうが,私はアナジャコに一番近いと予想した。分子系統解析の結果からは,両者は同じ属(Macrogebia 属)に含まれると結論される。
ただし,アナジャコとナルトアナジャコが分岐したのは,相当古い時代だろう。分岐年代の数値は示されていないが,古第三紀のあたりだろうか?イシアナジャコになると,中生代(白亜紀)にさかのぼるかもしれない。それらを全部ひとからげにして Upogebia と呼ぶ時代はもう過ぎている。
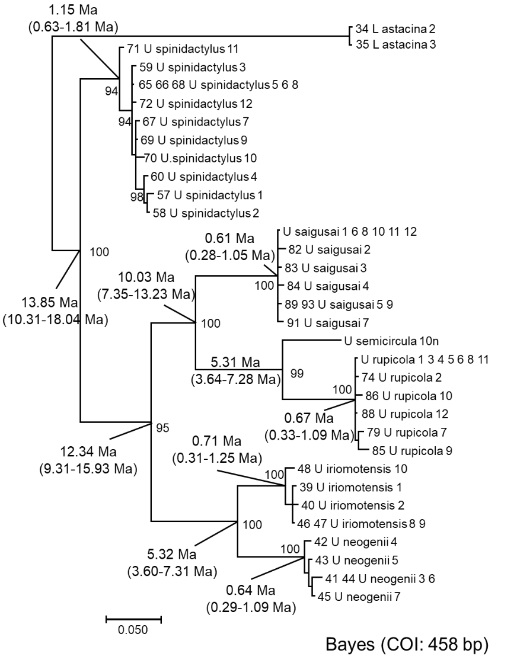
図 4.イシアナジャコ属(Stonogebia)の類縁関係と分岐年代。
イシアナジャコは,図 4 だけの内容から判断すると,spinidactylus と他の 5種類の含まれるグループの 2 つの属に分割できる。しかし,形態だけで見ると,spinidactylus が他のイシアナジャコよりも変わっているという感じはしない。rupicola は沖縄本島に広く分布しているイシアナジャコである。沖縄本島に分布しているのは rupicola だけ。
西表島に分布するイシアナジャコは 5 種類。neogenii は私がつけた種名。neogenii は形態的には iriomotensis と区別ができない。
直感を重視する分類学者に相談しても,形態学的に区別できない neogeniiなど,存在を絶対に認めようとはしない。結局,私たちの方で動物名(学名と和名)をつけるしか道はない。和名の公表は少し後で行うだろう。
また,saigusai は私がつけた名称ではない。spinosissima(棘が多い)に変更を考えている。形態学的には spinossima が一番違って見えるが,分子系統解析の結果は spinidactylus が一番離れていた。
琉球弧が存在したのは 5,000 万年以上前だが,西表島が現在の場所に顔を出したのは 1,400 万年前という説がある。偶然かもしれないが,spinidactylus が他の 5 種類のイシアナジャコと別れた時期とちょうど一致している。最初に西表島の河口に流れ着いたイシアナジャコは何という種類だろうか?また,spinidactylus の方は,全然種分化が起きなかったのに,もう片方のグループでは 5 種類のイシアナジャコが生まれている。どうしてなのか?
ひとつの考え方としては,spinidactylus は,1,400 万年前に南方の島で種分化を起こし,長い間その付近の島々で生活した後,割と最近になって西表島に流れ着いた問可能性もある。ただし,そう考える確かな証拠はない。
さらに,semicircula は,イシアナジャコの形態学を研究して博士号を取得した学生が命名した。私は何度も双眼実体顕微鏡で観察したが,他の種類と十分に区別できなかった。和名はあると思う。イシアナジャコの中で semicirculaの分布は局地的で,船浦の海岸に多く棲んでいる。semicircula はこれから西表島の西側全域に分布を広げて行くかもしれない。
お断り:図 4 には 1 か所間違いがある。U. rupicola の分岐年代が 0.67 Ma となっているが,U. semicircula と U. rupicola の分岐年代の誤り。また,アナジャコ類全般の分子系統解析の結果(図 3)は,イシアナジャコは他の属から独立していることを示すのではなく,イシアナジャコ類の中から泥や砂に穴を掘るグループ(属)が分化していったことを強く示唆しているだろう。その意味では,私がイシアナジャコ属(Stonogebia)と呼ぶのは間違っている可能性が高い。アナジャコ類の系統分類に関しては,もう一度分子系統解析の結果を整理して原著論文の形で世に出したいと思う。

図 5.イシアナジャコ類の生息場所。イシアナジャコは,琉球列島で 6 種類記録されている。西表島で 5 種類(a から e),沖縄本島で 1 種類(j と k)。分子系統解析の結果は,イシアナジャコ属の名称を支持していない。ナジャコ類の生息場所は,砂泥岩から泥に移行したことが予想される。
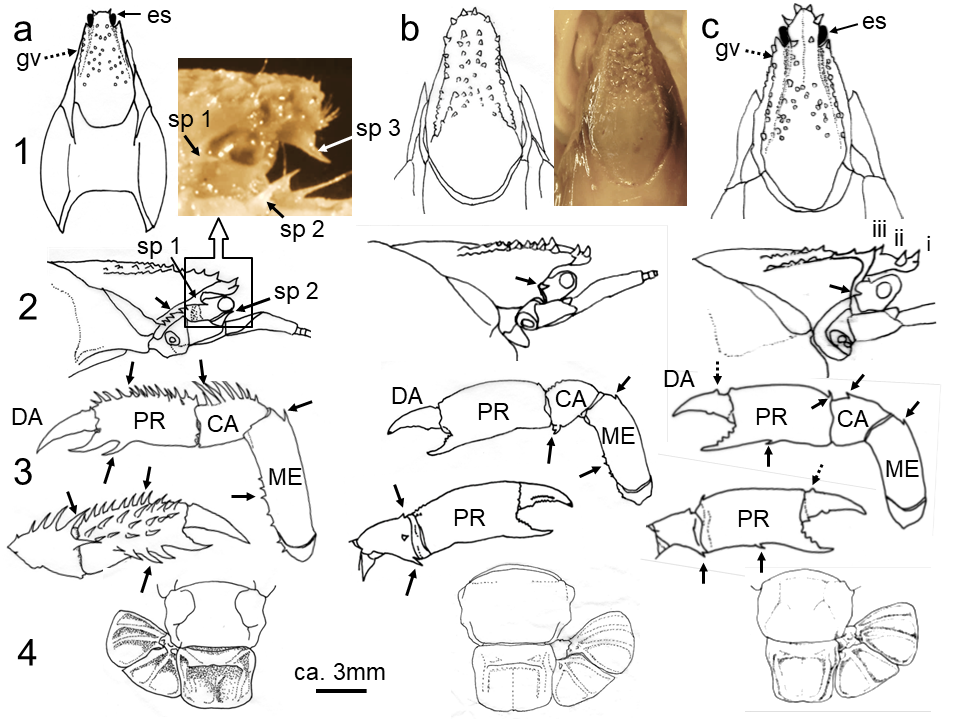
図 6.イシアナジャコ類の形態。a: spinosissima; b: semicircula; c: spinidactylusi。スケッチで見ると,各種の形態は異なるように見えるが,実際には双眼実体顕微鏡で見ても spinosissima 以外は容易に判別できない。故意にやっている訳ではないが,結果(スケッチ)は作為的になってしまうのだろう。
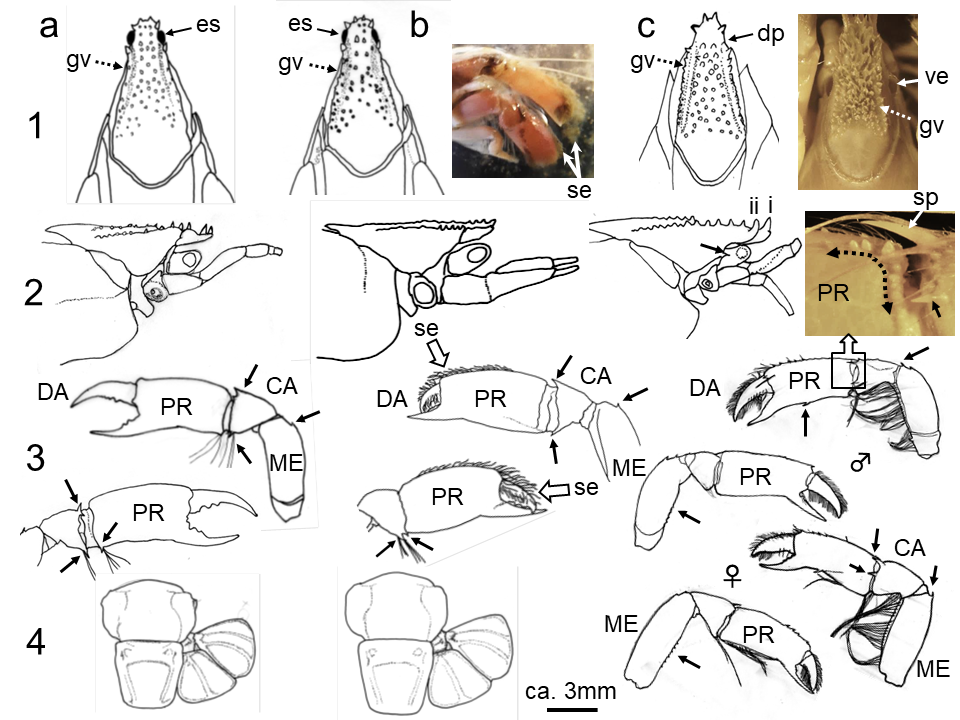
図 7.イシアナジャコ類の形態。a と b: iriomotensis か neogenii; c: rupicola。見かけは大きく違うといっても,収斂進化が進めば,形態で種を分けるのは難しいことがわかる(私の意見)。そもそも内生動物にとって,雌雄は判別されているのだろうか?「形態種」は意味がないかもしれない。
<検証可能な系統分類学に基づく人材育成>
大学には研究・教育の一環として研究室(laboratory)があり,そこで大学院生を中心に研究・教育が行われている。YouTubeで放映されている「コンバット」で言えば,私が軍曹で,博士課程学生が一等兵,修士課程学生や学部生が二等兵といったところだろうか・・・。しかし,私の方は,サンダース軍曹に比べ判断力・決断力は比較にならないほどレベルが低い。早い話,有能ではない指揮官が運営している組織(分隊)である。
もう20何年か前のことだが,研究室に生物の分類や形態を研究したいという女子学生が来た。偶然ではあるが,私はその前年に西表島でイシアナジャコ(図2と図5)を採集していた。4年次の課題研究ということもあり,形態の記載と系統学的な位置づけができれば,課題研究として「優」の認定ができる。・・・ということで,イシアナジャコの分類はどうかと声をかけたら,ぜひ研究してみたいという返事だった。この学生は,分子系統の研究はしたくなかったようだ。
この学生が入ってきたころ,私はどんな研究をしていたのかよく覚えていない。内分泌かく乱物質のプロジェクトが始まったころと思う。笠岡湾に棲むアナジャコ(Upogebia major)の腹肢に現れる形態異常と原因を調べたり,高知市春野町にある甲殿川(こうどのがわ)の河口や,広島県廿日市市の河口(川の名前は不明)に行って,干潟の泥に含まれる有機物の量とヒメアナジャコ個体群の成長の関係を比較する作業をしていたかもしれない。要するに,干潟に生息するアナジャコ類の生態や行動の研究を行っており,アナジャコを全く知らないという訳ではなかった。しかし,イシアナジャコに関しては,まだ素人の域にあった。砂泥岩に棲むイシアナジャコについては全然記録がなく,半ば言い訳にはなるが,知らないのも当然であった。
学生を指導するに当たって,十脚甲殻類の分類について専門的な知識を供与できる人はいないか,(現在の)共同研究者である知人に尋ね,ある人物を推薦してもらった。実は私の方は,この人物を知らない訳ではなかった。イシアナジャコを始めるずっと前に,私にところに電話がかかってきた。詳細は忘れたが,何でも教員の公募があって,私に推薦人になれという依頼だったと思う。私の方は,その人物については全く知らない。突然推薦人と言われても,「はあ?」としか答えようがなかった。
推薦状にもサインした覚えはないが,人事委員会の場には出されたのだろう。それを知った臨海実験所の所長なる者が,私に激しく怒ってきた。右も左もわからない若造に推薦人を依頼する者はどうかしているだろうが,逆に私が推薦人になったと言って怒り狂う所長なる者もどうかしていると思う。何かにつけて,ものすごい勘ぐり深い組織であったことは間違いない。大学というのはそんな人たちの集合体である。外には大きく出ないが,内側では激しいトラブルが絶えない。
イシアナジャコの方は,その人物に教えを乞うことにした。最初のうちは上手くいっていたようだが,次第に雲行きが怪しくなってきた。最初の論文は,図は少なくとも学生が書いている。本文(英語)については不明だが,学生が書いた原稿をその人物が手直しをしたかもしれない。この程度の貢献度ならば,研究者を育てるという観点から,学生がファースト・オーサーになるのが普通である。ところが,自分がファースト・オーサーになり,学生が2番目,私の名前が3番目という順序だった。記載したイシアナジャコには,私の名前がついていたが,私としては非常に迷惑な話であった。人の名前など入れずに,種(species)の特徴を示す(例えばspinicheles)とか,生息場所(habitat)の特徴を示す(例えばtropicusやiriomotei)とかできないものなのか?しかも,2つ目の論文では,およそ琉球弧に分布するとは思えない種類に突然変更した。やることが無茶苦茶である。検証などは頭になく,独断(神の意志)が先行してしまうのだろう。
2つ目からの論文では,学生が図を書き,本文も書いた。原稿の段階では,学生がファースト・オーサーになり,その人物がセカンド・オーサーになっていたと思う。私の名前も3番目には入っていただろうが,投稿する段階で削除された。そして,自分をファースト・オーサーにして,学生をセカンド・オーサーにして,論文をパブリッシュしてしまった。さらに次の論文も同様に自分がファースト・オーサー,学生をセカンド・オーサーにして論文をパブリッシュした。
このような論文の出し方には大きな問題がある。まず,学生本人が自分で手を動かして作成した論文にも拘らず,セカンド・オーサーにしなれないこと。その人物が指導したといっても,学生自身はその人物の研究室に行ったのは,5年間を通してせいぜい4~5回だったろう。形態観察や論文作成は,すべて私の研究室で行った。
だから,例の人物に対しては,共著者として名前を連ねることはおろか,謝辞(acknowledgments)の中でevery facilityを提供したと書くぐらいで十分ではないか。私の貢献度が少なかったのであれば,私も共著者から外して謝辞に入れてもよい。このようなケースの場合には,学生が単独の著者として論文をパブリッシュすることも可能である。しかし,サンプルを採集しているのは私なので,セカンド・オーサーとして共著者に加わるのがよいと思われる。
博士号を取得する際には,博士論文は本人が書く必要はあるが,それとは別に参考論文の提出が求められる。ただし,参考論文の取り扱いには規定がない。・・・なので,私は日本語で書かれた博士論文などよりも,英語で書かれた参考論文の方が,審査のためにはずっと大事だと考えていた。(大学では,大事なところに規定がなかったりする。逆に,社会がうるさい案件については,やたらと奇妙な規定があったりする。)審査の段階で,自分が指導した学生がよその研究室で丁稚奉公をしたような印象を与えると,研究指導上の無能さを指摘されることもある。そういう意見が大勢を占めれば,結果として博士号(doctoral degree)を取得できない可能性もでてくる。
私の力では,この人物の強欲さを止めることはできなかった。学生の方は,アナジャコのオスに見られる性的2型(sexual dimorphism)を研究し,びっくりするほど早く原著論文を書いた。博士課程修了とともに博士号を取得し,ある公的機関に就職した。この人は大学教員になれば,研究指導力は私より高かったかもしれない。
なお,性的2型に関しては,内分泌かく乱物質との関連で研究が行われ,環境汚染との関連性は低く,雌雄両性時代の名残(個体変異)という結論で決着した。
例の人物には,さらにひどい仕打ちを受けたが,もうこれ以上は書きたくない。学生が研究室を去った後に,私がこの研究を引き継いだ。例の人物について,博物館等に問い合わせたことがあった。数か所から「あの人にはとんでもない目に会わされた。十分に気をつけた方がよい。」との回答があった。いつも自分のことだけが大事な人なのだろう。今はどこかに雲隠れしているのかもしれない。
分類学者の中には,太刀の悪い習慣を持った人がいる。(変な習慣を持つ人は,すべての分野にわたって多い。)そのひとつが,採集した標本を自分の所有物にして,独断的な記載をし,論文を発表すること。偶然に希少な標本を捕まえたという例もあるが,希少かどうかはある程度動物を見慣れていれば,おおよその判断をつけられる。だから,自分の判断でよいか分類学者に確かめる例が多いかと思う。
分類学者は標本を手に入れると,新種もしくは希少種であれば,採集者を無視して自分の名前だけで論文を発表するケースも多い。権威ある人に対しては平身低頭し,論文は共著で発表する。自分より権威の劣る人々に対しては,標本を採集した人のことは全く無視する。標本を調べたものに単著で論文を書く権利があるなどと,矛盾したことを平気で言いふらす。しかも,種や属の同定は,独断(神がかった判断)で行われるために,著しく客観性を欠く。
もうひとつの問題は,種名に人名をつけている例が多いことである。生物の記載の仕方には一定のルールがある。カール・フォン・リンネ(1707‒ 1778)が提唱した2命名法である。この方法では,最初に属名(genus),次に種名(species)を併記する。両者は通例イタリックスで表示される。(筆記体の時は属名と種名に下線を引く。)
種名や属名に,人名を用いるというのは大昔の話である。例えば,丘浅次郎(明治時代の官僚的文化人,形態学者)は自分の名前をもじって,カンテンコケムシ(これは和名)に Asajirella gelatinosa Oka, 1891という学名をつけた。こんなやり方は,あまりにも古すぎる。
種名に人名がついた分類学の論文を見ると,まるで平安貴族が蹴鞠遊びかカルタ取りをしているようなイメージを与える。もちろん、そういう遊びごとを馬鹿にしている訳ではない。昆虫や甲殻類の図鑑等を用いて,名前当てクイズを行う人たち(一部の生態学も含む)は,それで十分満足かもしれない。この種を始めて記載した(命名者)のは,実はあの人だった。・・・みたいに尊敬されるかもしれない。
一方,人名が種名や属名の中に入ると,論文を書くときに,いつの間にか命名者が顔を出してくる感覚に襲われる。特にトラブルがあった人物に対しては,忌々しい思い出がよみがえってきて,(キーボードの)キーを打つ手がとまる。場合によっては,命名者をたたえる論文になる可能性もある。人名は記号の一種だと主張する人がいる。そう自分に言い聞かせて使ってみると,やはり特定の人物(命名した者)のイメージが付きまとい,論文は命名者をたたえるような雰囲気に変わって行く。自分から出かけて行って採集した訳でもなく,単にお経を唱えただけ(つまり命名しただけのこと)で,何でそんなに尊敬されるのか,私には全く理解できない。
蹴鞠遊びやカルタ取りがお好きなノスタルジア分類学者たちに言いたい。学名をつける際には,種名の後に命名者の名がつく訳だから,それ十分ではないか。種名や属名の中にまで自分の名誉を付与する必要はない・・・と。分類学者の悪習は,野鳥では比較的少ないように思えるが,昆虫(特に甲虫類)と十脚甲殻類では,うんざりするほど多くの人名が出てくる。現代生物学は,ノスタルジア分類学から決別しつつある。種(species)や属(genus)の名称も,2命名法というルールは変更しないが,人名は徐々に消える運命にある。
もう一度,例の人物の話に戻る。例の人物が執筆した十脚甲殻類の分類に関する本がある。オランダかドイツの博物館の所蔵標本を引っ張り出して,標本のスケッチを並べている。アナジャコ類に関しては,ほぼすべての標本が同じ属(genus)になっていたように記憶している。アナジャコやスナモグリの起源は不明であるが,ともに単系統(monophyly)であることは間違いないように思える。カニやヤドカリと同じような時期,中生代のジュラ紀に出現した感じがする。
もっとも,カニとヤドカリの起源は多系統(polyphyly)かもしれない。いずれにしても,中生代に誕生した生物が,全部同じ属というのは考えにくい。いやいや,新生代第四紀に入って50万年から100万年前に一斉に種分化(speciation)が起きたという形而上学的説明(言い訳)も成り立つ。しかし,海岸に堆積したサンゴ礫やサンゴ砂に深い穴を掘る性質や,砂泥岩に巣穴を作る性質が数百万年で進化した可能性はまずない。数千万年か数億年の単位で可能になる変化だろう。
直感的分類学は,形而上学の一種である。物の判断,例えば個体変異・種レベルの違い・属レベルの違いの判断は,いわゆる権威者からの「ツルの一声」である。ノスタルジア分類学者の主張する「証拠」は,客観性が完全に欠如していて,権威者の独断のみに頼っている。
例の人物が私にあてた手紙には,自分は世界的権威であることが書かれていた。普通は自分からそんなことは書かないので,相当いかれた男なのだろう。権威者の言うことならば,下々の人間はすべて信じるとでも思っているのだろうか?
この20年間に分子レベルでの系統解析を行う分野(分子系統学)が大きな進歩を遂げた。ノスタルジア分類学の知見と一致するケースもあるが,大きく異なるケースも多い。特に十脚甲殻類に関しては,穴掘り動物ということもあり,形態の収斂(convergence)が進んでいる。形態的には全く同じ種のように見えても,分子系統解析をしてみると,実は別種のこともある。場合によっては別属とみなす方がよい例も頻繁に出ている。同様なケースは今後増加するに違いない。
分子系統解析の結果と権威者の行う主観的分類の結果が大きく異なる場合には,客観性の高さを考慮して,分子系統解析の結果を優先するだろう。分子系統解析の結果,和名も含めて新しい学名を置くということが必要になることもある。大事なことは,新しい種名や属名の設定は,分子系統解析をやっている方の研究者が行うこと。もちろん,形態はよく比較してから実行するべきである。直感的・ノスタルジア分類学者に相談すると,上のいくつかのパラグラフで述べたような問題が再燃する。今まで長い文章を書いてきた理由はここにある。
例の人物を含めて,リンネ分類学から一歩も出なかった人たちは,悔しいだろうが,人間と学問が古すぎることを自覚すべきである。Dworschakだったか Ngだったか,例の人物に味方する人たちのセンスでは,現代生物学の流れについてゆけない。
みんなもうお墓に入るころだろうから,黙って見送りたい。カール・フォン・リンネがお墓の中で待っているだろうから,DworschakとかNgとか,Manningとかいう人たちを誘って,リンネを囲み,みんなで一杯やりながら昔話(自慢話)に花を咲かせたらよい。
必ずしも若い人たちということではないが,客観的な手法を用いて動物の分類,形態学,系統学の研究をめざしている人たちに道を譲ってあげたらどうだろうか? 若い人たちだっていつかは同じようにして新しい波に押されて消えて行く。みんな同じなのである。自分だけが馬鹿にされている,などという被害妄想を持たないことだろう,それに自分は権威者だなどと慢心しないことが肝心である。

図 8.イシアナジャコの生息場所。
イシアナジャコ類は,内湾(inlet)の海岸と河口域(マングローブ)の干潟に露出している砂泥岩に巣穴を作って棲む。砂泥岩は,砂の粒度より石や岩の硬さが問題だろう。西表島の砂岩は,非常に硬いものから,図 8 のようにノミやトンカチで容易に崩せる柔らかいものまで幅広い。硬い砂泥岩には住んでいない。スケールは割りばし。
巣穴には入り口が 2 つある(図 5 の b を参照)。巣穴の出口は狭くなっていて,ここからイシアナジャコが出入りする可能性は低い。また,干潮時の観察しかないが,巣穴を出て石や岩の上を歩いているイシアナジャコも見たことがない。イシアナジャコは,おそらく一生を巣穴の中で過ごすのだろう。
では,生殖時期になって雌雄で交尾は行われないのだろうか?私は,雌雄が出会って交尾しなくても,産卵(受精)は可能ではないかと考えている。イシアナジャコは,ひとつの砂泥岩にたくさん集まってそれぞれの巣穴を作っている。大潮の満潮時に成熟したオスが一斉に精子を放出すれば,メスの生殖口には届くだろう。イシアナジャコは,異性を認識できなくても種の維持ができている可能性も視野に入れておきたい。

図 9.イシアナジャコの棲む砂泥岩。岩の上の層は砂岩のようだが,泥浜に接する層は,水を含んで柔らかい。生息している種については,肉眼ではわからないが,spinidactylus, iriomotensis, neogenii と思われる。削ったところは灰色(粘土質?)。個体数は非常に多い。崎山湾の奥で撮影(2007 年 5 月)。

図 10.イシアナジャコの棲む砂泥岩。岩の上の層は砂岩のようだが,泥浜に接する層は,もう少し泥が混じって柔らかい。生息している種については,肉眼ではわからないが,spinidactylus, iriomotensis, neogenii と思われる。崩した部分は灰色(粘土質?)。個体数は非常に多い。崎山湾で撮影。

図 11.イシアナジャコの棲む砂泥岩。大潮の干潮時(崎山湾で撮影)。堆積した泥の表面から 30cm ほど上までの範囲の砂泥岩の層に棲んでいる。一方,泥の中には小さなスナモグリ(Callianassa vouvieri)がいる。岩や石の隙間にはカニ類が多く,巣穴を出たらすぐに捕食される。

図 12.泥干潟に崩れ落ちた砂泥岩。おそらく数年前に崩れ落ちたのだろう。イシアナジャコがいつ砂泥岩に入り込むのか不明だが,5~6 年はかかるような気がする。しかし,時間がたてば砂泥岩は泥の下に埋もれてしまう。イシアナジャコは、いつまでも同じ砂泥岩の中にはいられない。

図 13.崖から落ちた砂岩。右上に向かって突き出ている地層は何だろうか?すぐ後ろに砂浜が見えるが,岩があるところは潮上帯(supratidal zone)になり,イシアナジャコの巣穴はない。層によっては,スナモグリやアナジャコの化石が出ると思う。崖の植物:アダン,トベラ,シャリンバイが多い。

図 14.崎山湾の泥干潟(大潮の干潮時)。このあたりの砂泥岩は最近崖から落ちたものらしく,イシアナジャコの巣穴は少ない。手前のタイドプールにはスズメダイ科の魚が多い。石を取り除きながら網ですくえばいくらでも採集できる。・・・が,そんなことをしているとすぐに潮が上げてくる。

図 15.崎山湾の泥干潟(大潮の干潮時)。干潟の茶色の泥は,写真が古いのではなく(フィルムを使っている時代ではない),地層の色が出ている。正面にヤエヤマヒルギの群落が見える。かつての崎山集落はヒルギ林の奥にあったのだろう。崎山は 1948 年に廃村になり,今はその面影も残っていない。
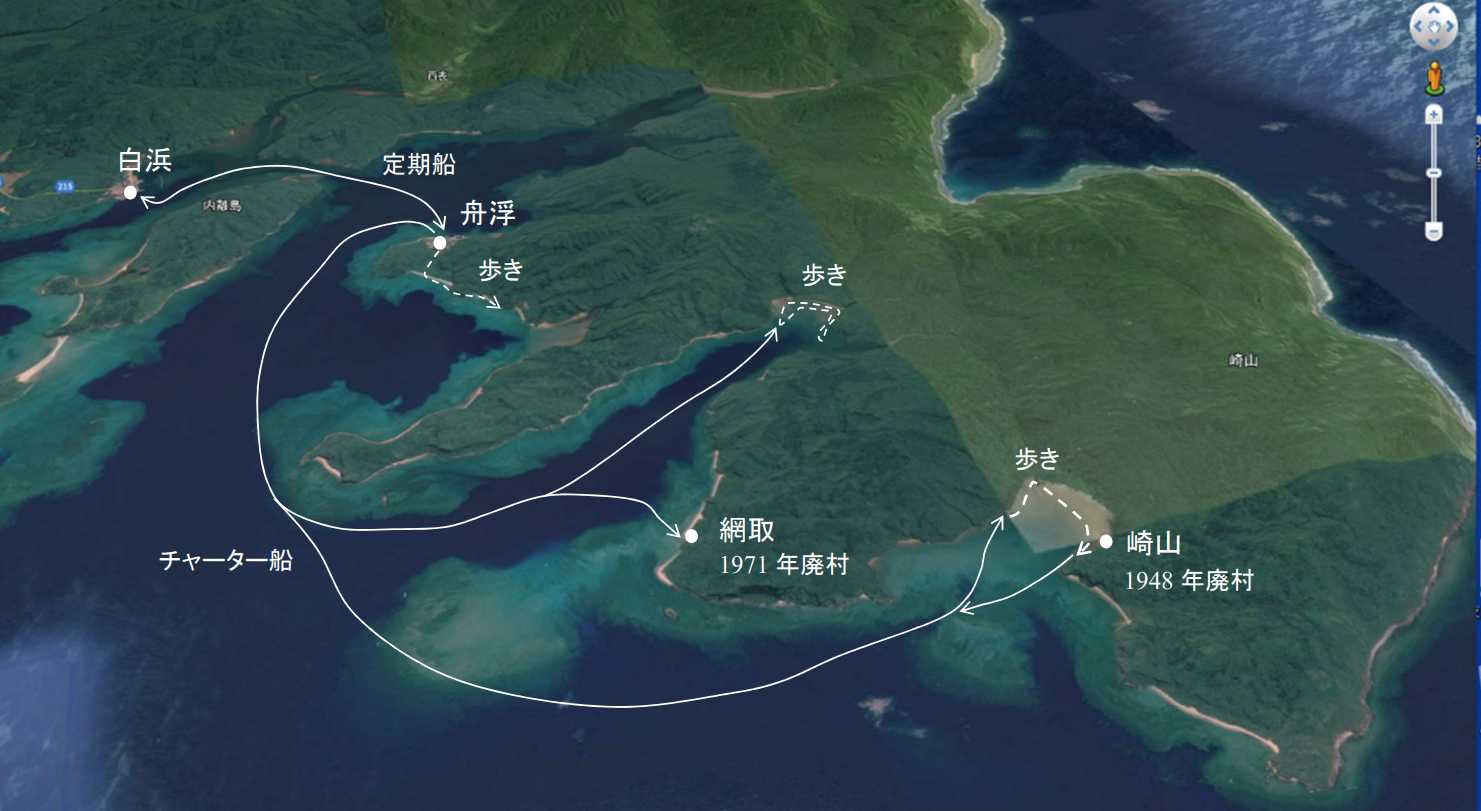
図 16.西表島・舟浮から奥の湾の泥干潟。白浜から舟浮までは定期船があって毎日 4 回ほど往復している。白浜から舟浮の海岸や河口には砂泥岩が広く分布し,イシアナジャコは非常に多い。舟浮からイダの浜を通って海岸沿いを歩くと,狭い泥干潟がある。網取・崎山方面は,船(ボート)をチャーターする。調査で行くなら舟浮海運の元船長に頼めばよい。西表西部の地形や海の様子を一番よく知っている。レジャーボートを持っている観光客目当てのガイドはこういう案内には不向き。西表島の自然をよく知らない。一方,元船長はもう相当な年だろうから,行ってくれるかどうか?今の舟浮海運の船長は,元船長のご子息と思われるが,こちらは定期船の運航で忙しい。網取(あみとり)の海岸には,柔らかい砂泥岩はなく,イシアナジャコはいない。一方,網取湾の奥には泥干潟が広がり,イシアナジャコは多い。昔は世捨て人の爺さんがひとり,犬と住んでいたが,今はどうなったか?一度会ったことがある。崎山(さきやま)湾には広大な泥干潟があり,湾全体にトゲスナモグリが作った砂の塚がある。崎山湾にもイシアナジャコは多い。

図 17.網取湾や崎山湾に行くチャーター船。船を操作するのは舟浮海運の元船長。小さいころから舟浮の集落で育ったのだと思う。海をよく知っている。浜でボートを降りた後は,ずっと一人で海岸沿いをぶらぶら歩いた。夕方になるとボートが来て,白浜まで戻った。2007 年に撮影。

図 18.網取小・中学校の卒業・終業式の記念写真。(「竹富町史だより」(竹富町史編集室)第 13 号の表紙を転載)。1969 年(昭和 44 年)撮影。崎山集落は,すでに 1948 年に廃村になっているので,小・中学校の卒業記念写真はないかもしれない。網取小・中学校は,1898 年(明治 31 年)に大川尋常小学校崎山分教場として創立された。1898 年というと,本州では伊藤内閣,大隈内閣,山形内閣と目まぐるしく変わり,市町村立学校に学校医制度が導入された。前年度である 1897 年には,帝国大學が東京帝国大學に改称され,2 番目の京都帝国大學が設置された。
網取集落は 1971 年に廃村になった。この写真(図 18)は,廃村の 2 年前(1969 年)に撮影されている。卒業生(前列右側の 3 名の女子生徒か?)は,1969 年で 12 歳か 15 歳だろうから,1954 年か 1957 年生まれだろう。とすると,2024 年(令和 6 年)には 67 歳か 70 歳前後で,皆まだ元気で暮らしているに違いない。今どこで何をしているのだろうか?また,1969 年の在校生は,今(2024)は 60 歳から 69 歳ぐらいになっているだろう。網取集落が廃村になって波照間島に戻ったのだろうか?それとも,石垣島に移住したのだろうか?あるいは,西表島のどこかの集落で生活しているのだろうか?

図 19.崎山湾から白浜へ。西表島の西海岸には垂直に近い崖が連なっている。最上段の地層はほぼ全域にわたり堆積岩(八重山層?西表層?)である。西表島の砂岩は脆い上に,西表島全体が西の方向に向かって沈みつつある。西側の海岸では,海岸に張り出した地層が頻繁に崩落し,垂直に近い崖ができるのだろう。島の中央に道路を作っても,頻繁に起きるがけ崩れのために,通行止め箇所が続出するに違いない。建設を中止して正解だった。
西表島の西側の海岸とは逆に,東側の海岸は多少浮き上がった状態になり,陸上から流出した泥が河口や海岸に堆積して広大な泥干潟が現れる。ただ,広大な干潟は栄養も豊富だが,生物にとって隠れ場所がない。そのため東側の泥干潟に棲む生物の種類は,西側に比べて圧倒的に少ない。