
1.Introduction
十脚甲殻類(decapod crustaceans)には,水産資源という面から重要な種類がたくさん含まれるが,地球上に無限にあるという資源ではない。現代の優れた技術を使えば,瞬く間に一網打尽にされ,ある時点を境に漁獲量は急激に減少するだろう。昔はいっぱい獲れたが,今は漁獲量が昔の100分の1以下になってしまった,なんていう話はざらにある。漁獲量を規制したところで,すぐに抜け道ができる。また,厳しく資源管理をしても,よその国が漁場に侵入して好き放題に捕って行かれたらたまったものではない。
水産資源を確保するために養殖(aquaculture)という手段がある。植物の場合には制御する環境要因が比較的似ているので,養殖(植物の場合には人工飼育,または農業というのだろうか?)は,技術が確立している。農業の方は,いつ何をすればよいかを守れば,最大収穫量を得ることができる。
一方,動物,特に水産無脊椎動物の場合には,海の中のどんな環境で育っているのか,また何を食べているのか,よくわかっていない種類が多いのではないだろうか?食べるエサの種類や大きさも,発育段階に応じて異なっている種類も多いだろう。こんな事情もあって,海産動物の養殖は,一般に難度が高い。一番の苦労は,ほとんど毎日動物につききりで世話をしなくてはならないことだ。農業の方は,決まった時期に決まった量の肥料や農薬を施せば結果が保証されるのに比べ,養殖業の場合には,水質がわずかに悪化すれば,水槽で飼育している魚介類が全滅することがある。とにかく,四六時中動物の状態を見張っていなければならない。これはものすごく忍耐力のいる仕事であり,その苦労たるや飼育に携わった人たちでないとわからない。
私は,アカテガニの産卵の太陰リズム(lunar rhythm)の発現機構を研究するために,アカテガニ(Sesarma haematocheir)を使った。4月中旬から下旬にかけて,志摩(三重県)や笠岡(岡山県)の河口の土手に行き,冬眠しているアカテガニを採集し,実験室(恒温室)に持ち帰った。こんなところでうまくメスは抱卵してくれるのか疑問に思いながら,隠れ場所や脱皮場所を準備し,毎日パンとか昆虫をエサとして与えた。抱卵すると,底に水を張った衣装ケースを置いて,板切れを利用し,ゾエア幼生が孵化するまで抱卵メスが隠れていられるアパートを(衣装ケースの中に)設置した。豆電球を使った人工月光(artificial moonlight, 0.5 lux)のonとoffの時刻は,タイマーを使って毎日セットし直した(その頃プログラムはなかった。)実験期間は4月末から確か8月いっぱいぐらいだった。毎日の仕事量はそう多い訳ではなかったが,半年近くにわたり,一日の休みもなくカニの世話をするのは気苦労が多かった。
医歯薬系や工学系の分野では,実験動物としてネズミとかウサギとか,飼育業者から購入して実験に使用しているのだろう。これらの学部では,研究はネズミとかウサギを使ってやるものだという傲慢さが蔓延しているようだ。一方,理学部生物学科では,研究対象とする動物を広く自然界に求めてきた。昔は,ウニ,ホヤ,ナメクジウオ,カブトガニなど,多くの海産動物を飼育して研究に使った。また,農学部では,多くの種類の昆虫類を使った研究が行われてきた。
自然に生きる動物を研究で使用する場合には,対象とする動物を飼育する必要がある。昆虫を使う場合には,研究室の中に低温恒温機,もしくは恒温室があればよいが,海産動物は,大量に生の海水(fresh water)を使う場合が多く,研究室では長期の飼育は難しい面がある(不可能という訳ではない)。手っ取り早いのは,各地の大学の臨海実験所(marine laboratory)を利用することである。臨海実験所や水産研究所であれば,生の海水は自由に使える。昭和のころまでは,海産無脊椎動物を研究に使う人が多く,臨海実験所は大盛況であった。
ところが,昭和も終わりごろになると,臨海実験所の利用者は急に減少した。原因は,海産動物を使って研究する人が減少したことだろう。研究者の減少に伴って,臨海実習のニーズも少なくなった。今では,全国各地にある理学部附属の臨海実験所は利用者もなく,閑散としている。理学部附属の臨海実験所の利用者数が大幅に減少していることは,すでに文部科学省の耳にも入っていると思う。改善が試みられているが,根本的な問題が解決できていないために,改善の効果は期待できない。
利用者数の減少は,何も臨海実験所に限ったことではなく,文部科学省や各県が管理する教育研修施設においても同様である。日本では,臨海実験所とか研修施設は,子供たちや学生を連れてきて教育する場だという意識が強すぎるのではないか?修学旅行ですら,未だにそんな意識で実施している学校が多いと思う。
子供たちは,野外に出て遊びたい。しかし,野外には危険な場所も少なくない。野外研修(登山とか水泳の訓練も含む)にはリスクが伴う。子供本人も保護者もそのことをよく理解していないので,事故は多発する。事故が起きれば,当然ながら管理責任が問われる。判断力の悪い校長がいれば,原因究明はそっちのけで,事故を隠蔽する者もいるだろうし,逆に一方的に指導者に責任を押し付けて一件落着を図る者もいる。いずれにしても,わざわざリスクを冒してまで野外研修を実施しようと思う人たち自体も,相当減少しているように思う。
とは言え,野外研修(要するに「訓練する」ということ)は,子供たちの人格形成にとっては重要である。人格形成を考えるのであれば,「二十四の瞳」的教育がよいだろう。しかし,現在は「大学入試センター試験」的教育が主流になっている。私は,道徳は学校で教えるのではなく,保護者が教えるべきだと思っている。家族でレストランに行ってマナーを議論形式で教えたり,道路横断の危険個所を教えたりできる。キャンプに行ってゴミ問題のことを話し合うこともできる。親子でゲームセンターに行ってもよい。いろいろな野外研修ができるだろう。ただ,パチスロはまずいぞ・・・。
もうひとつ,臨海実験所が利用されなくなった理由として居住性の問題がある。各地の臨海実験所や研修施設には,「二十四の瞳」的教育精神があふれている。ホテルや民宿に来るような気分で来てはいけないし,家族で来ることも厳禁である。臨海実験所では,昔ながらの生活様式が維持されている。一言でいえば,とにかく窮屈である。実験所は,研究するための施設だから仕方ないと言うだろうが,それなら近くのビジネスホテルにでも泊まった方がよい,となる。
現在私がよく利用する実験所は,琉球大学熱帯圏研究センター・西表研究施設である。場所は西表島の上原にある。私の場合,夜出て行って調査をすることが多いので,民宿はちょっと無理。宿泊棟は個室でシャワーもあり,エアコンもついている。小さな冷蔵庫もあるので,ビールを冷やしておける。標本の整理も部屋の中でやれる(公には禁止)。朝食や夕食も,割と好きな時間に食べられる。雨が降れば,部屋で記事を書いて過ごせる。眠くなれば,自由に寝られる。
施設の利用は,干潟生物の観察という利用目的にすれば,個人でも家族でも可能だろう。私は西表島では,いつも一人で活動している。民宿やホテルに泊まるよりも,研究施設に泊まる方が,メリットは大きい。琉大熱研の西表研究施設と瀬底臨海実験所(名称は変更になっている)には,私と同じような利用者が多いと思う。海外の臨海実験所も,私のような使い方をしている研究者は多いと思う。
日本の多くの大学には,海洋科学(海洋物理学や海洋生物学)に関する授業科目はない。しかし,日本の地理的特性を考慮すれば,理学部,農学部,教育学部,工学部には,海洋生物学の授業科目はあった方がよい。授業科目にない以上,海洋生物学を学ぶためには,まず海岸(seashore)に出て遊ぶことから始まる。遊ぶ中から学問を立ち上げて行けるだろう。ただし,野外にはいたるところに危険が待ち構えている。くれぐれも身の安全には気を付けられたい。
自然を相手に「遊ぶ」などというとすごく怒る管理者が多い。大学を含めて学校現場には,未だに「二十四の瞳」的,「二宮尊徳」的教育精神が強く残っている。「二十四の瞳」的教育精神は,「面倒くさい」という理由で今の人たちには敬遠されるだろう。結果として,臨海実験所や各地の研修施設は衰退してしまった。
臨海実験所に関しては,今後どのような改善が期待されるだろうか?私は,見かけ上改善しても,理学部附属のままだと,依然として「二十四の瞳」的,あるいは「二宮尊徳」的教育精神が亡霊として残るだろうから,利用者は増えないと思う。
理学部から切り離して,(学校)教育学部附属センター(例えば,海洋教育実習センターと改称)し,初等学校や中等学校の教員を目指す人たちに利用してもらうのはどうだろうか?学校教育の教員研修にも利用しやすいだろう。SSHの実習にも大いに利用できる。教育学部にもそのぐらいの施設があってもおかしくない。
一方,施設の管理替え(既得権益の移譲)となると,大学の人たちは人間が変わる。しかし,環境何とかという学部を,丸ごとひとつ取り潰しにしたぐらいだから,頑張れば実現できる可能性は高い。
食卓に上がる前の十脚甲殻類を見るには,底引網漁(trawl fishing)をやっている漁船がつく港にある水産物直売所を訪れてみるとよい。一方,十脚甲殻類の系統と進化に関する研究をするならば,食卓に上がらない種類も重要だ。そのような十脚甲殻類を得るためには,船が着く漁協の桟橋に行く必要がある。
しかし,食卓に上がらない十脚甲殻類は,網を上げると船の中で選別され,海に捨てられる。そうされないためには,魚介類を入れる木箱(トロ箱と呼んでいる)を購入するという条件で桟橋まで持ってきてもらえばよい。ほとんどはガラクタ(主には貝殻と魚)しか入っていないが,10箱単位ぐらいで安く購入できる。トロ箱は,私が桟橋に行くまで漁協の冷蔵庫に保管してもらえるというメリットがある。
2.撮影と執筆の基本情報
<撮影場所> 境港水産物直売センター(鳥取県),沼津魚市場INO(静岡県)など。
<撮影と記事の執筆> 三枝誠行(NPO法人,生物多様性研究・教育プロジェクト常任理事)。<撮影機材> SONY RX10Ⅲなど。
<Key words> 動物の飼育,海岸で遊ぶ,十脚甲殻類,トロ箱,基幹構造,付属肢,胸部腹側のプレート,進化。
3.参考文献
Castro, P., and M.E. Huber (2005) Marine Biology (Fifth Edition) McGraw-Hill Higher Education.
Nybakken, J.W. (2001) Marine Biology: An Ecological Approach. Benjamin Cummings.
ガザミ種苗生産研究会(編)(1997)『ガザミ種苗生産技術の理論と実践』 日本栽培漁業協会
Saigusa, M. 1980. Entrainment of a semilunar rhythm by a simulated moonlight cycle in the terrestrial crab, Sesarma haematocheir. Oecologia 46: 38-44.
Saigusa, M. 1988. Entrainment of a tidal and semilunar rhythms by artificial moonlight cycles. Biological Bulletin 174: 126-138.
坪井栄(1952)『二十四の瞳』 光文社
山田真弓・西田誠・丸山工作(1971)『進化系統学』 裳華房

図 1.モニターの上でくつろぐぴよ吉(ブッポウソウ)。ぴよ吉は自由に部屋の中を飛び回り,目を合わせると飛んでくる。写真左にトイレがある。

図 2.モニターの上にとまるブッポウソウとベランダの手すりにとまるハシボソガラス。ガラス越しに距離は 70~80cm。2 匹で同じ方向を向いて体が固まっている。どっちも鳴き声は出さず。このカラスは首の背側に白い羽毛(の束)があるので個体識別ができる。私に対する警戒心は低い。

図 3.大山(鳥取県)における合宿研修。この時には新入生の合宿研修だったか,忘れてしまった。大山の頂上付近には雲がかかっている。

図 4.大山(だいせん)自然歴史館で大山の自然について説明を受ける学生。説明しているのは歴史館のスタッフ。私ではない。

図 5.文殊堂(大山)での灯火採集(LED の光では虫は来ないので注意)。この時には文部科学省からの経費を使ったように思う。宿泊した研修所(中国四国大学合同合宿研修所?)はもうない。いい研修所で,何度も利用した。大山や蒜山で合宿研修をやるときには、どこを利用したらよいのだろうか?

図 6.広島大学理学部附属臨海実験所における合宿研修。は学内の教育経費を使ったかもしれない。学生は遊びに出るときにはやたらと元気になる。

図 7.境港水産物直売センター(鳥取県)。昨年(2022)の 12 月 20 日ごろ。20 日までは人は少ないが,下旬になるとこの 10 倍ぐらいになる。岡山から行くとなると,12 月 20 日を過ぎると積雪で四十曲峠を越えられない時がある。12 月 10 日を過ぎると,毎日インターネットで鳥取県の積雪情報をチェックしている。境港水産物直売センターでは,日本海で獲れたベニズワイガニと輸入したタラバガニを買える。
合宿研修では大山とか蒜山に行くが,境港水産物直売センターの方が楽しくて,勉強になると思う。ただし,そんな研修が学部内でばれたら大事(おおごと)になる。大学には,食べられる生物を研究しろと言っているくせに,こういうところに研修に来ると激しく怒り出すチョンチョロゲな管理者が多い。そんな奴らが,コンプライアンスだ何だと騒いでいるのを見ると,大学というのは一体何をするところなのかと,不信感は増すばかりである。しかし,自分が管理者の立場に立ったら,同じようにやっていた可能性が高い。そう思うと,自分がちょっと嫌になってくる。

図 8.境港水産物直売センターで販売されているベニズワイガニ。私は建物中を 10 回以上回って,安くて新鮮な感じのするカニを購入する。5 匹で 5.500円ぐらい(壊れた個体を一匹おまけにもらう)ならば,妥当な値段だろう。販売所の店員は,身が入っているかを気にしているが,これは高く売るための口実である。ベニズワイ(ズワイガニも同じ種?)は,水深 500m−2,700m (1,000m−2,000m に多い)で採集される。
脱皮直後に殻(甲羅)が柔らかいのは当然だが,普通のカニと違って,脱皮後の殻は少し厚くなるだけで,手触りとしてはずっと柔らかいのだろう。脱皮直後の個体を除けば,どの個体も味は同じと思う。カニ味噌が多いかは,甲羅を触っただけではわからない。(店員も結構いい加減にやっている感じがする。)ベニズワイは,殻以外は全部食べられるのがすごい。ベニズワイはこれからますます値が上がるだろう。形にこだわらずに新鮮で安い個体を買ってたくさん食べておかれたい。

図 9.水産物直売センターの敷地内にある和食レストラン(海陽亭)のメニュー。半分ぐらいのメニューに十脚甲殻類(エビとカニ)が使われている。直売センターがごった返しているときには,境港駅の構内にある回転すし店(名前は忘れた)に行くのもよい。面白い具があって楽しく食べられる。
境港に行ってカニを買ったり,海鮮丼を食べたりするのもいいが,12 月 20 日あたりだと鳥取県と岡山県の県境にある四十曲峠(しじゅうまがりとうげ)にはいつ雪が降るかわからない。水産物直売センターであまりのんびりしてはいられない。15:30 ごろまでに峠を越える心づもりで帰路に就く。

図 10.ロブスター(Homarus americanus)。カナダ産と思う。頭部はザリガニによく似ているが,ゆでて食べるとすごくおいしい。ロブスターは,鋏脚にも身がしっかりと詰まっていて,すごくうまい。ザリガニ類は,ロブスターによく似ているので,日本のスーパーでは売れないかもしれない。
ザリガニはロブスターの祖先が 2 億年近く前に陸に上がって,世界中に広がったのだろう。ロブスターの祖先から進化した十脚甲殻類としては,ヤドカリ類,多くのアナエビ類(全部ではない),アナジャコ類,スナモグリ類が考えられる。カニ,カニダマシ,コシオリエビ,イセエビ,アカザエビは別系統のように思われる。これら 2 つの大きなグループの祖先は,三畳紀末に分かれて独自に進化してきたと思われる。

図 11.オキナワアナジャコ。西表島・古見で採集。マングローブとその周辺の非常に広い範囲(泥の中)を移動し,マングローブの縁に大きな塚を作る。もともとは河口の軟泥地に住んでいた古アナジャコの一種が,新生代に入ってマングローブ域の拡大に伴い,大型化したのだろう。オキナワアナジャコは,食べるところはほとんどない。しかも,泥臭いだろう。この個体は学術目的で採集したのであって,食べるために捕獲したのではない。

図 11.早朝の沼津魚市場(セリの始まる直前の風景)。2008 年 4 月。 仲買の人たちが集まりだすとそれぞれの区画でセリが始まる。セリの際に使用される言語は何を言っているか意味不明。昔(2008 年)はここまで入れてもらえたが,今は無理。また,ここに並んでいる魚介類を買えるのは仲買人のみ。

図 12.アカザエビ。エビではなくロブスターの仲間。10 年以上前から,駿河湾ではアカザエビは獲れていないと思う(乱獲)。食卓には出回らない。

図 13.ゾウリエビ(腹面)。腹部には身(筋肉)が詰まっていておいしい。胸部の腹側には,歩脚の間にプレートがある点に注目していただきたい。

図 14.タカアシガニ。小さいけど,10 年ぐらいは経っていそうだ。ゆでて食べてみたことはあるが,脚の殻が固く,厚い。身は少ない。味は今一つ。

図 15.タカアシガニの頭部と口器。頭部には複数の棘と第 2 触角が,口器では第 2 顎脚と第 3 顎脚が見える。頭部は鎧武者みたいな恰好をしている。

図 16.ミズヒキガニ。歩脚は 5 対。十脚甲殻類の歩脚(指節と掌節)はいろいろな形をしている。大きさからして食べるのは無理。味は不明。

図 17.市場の外で拾った十脚甲殻類。セリの後,かごやトロ箱を水で洗い流す時に捨てられるゴミに混じる。センジュエビやアナエビなどのお宝もある。
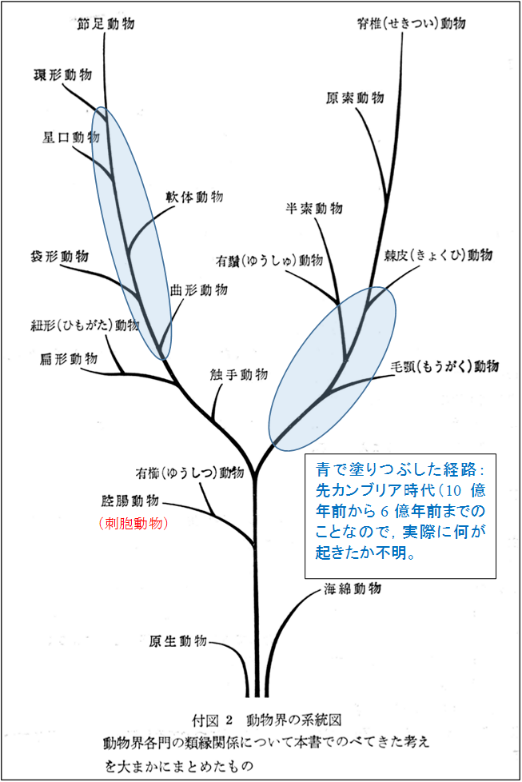
図 18.動物界の系統図(phylogeny)
動物の系統図に関しては,一度自分で書いた覚えがあるが,ファイルが見つからない。また書いている時間がないので,山田・西田・丸山(1971)進化系統学,裳華房より転載した。
この系統図には 2 つの大きな間違いがある。ひとつは,動物が刺胞動物(左側の図 18 では腔腸動物になっている)で 2 つに分かれてから,片方は節足動物に,もう片方は脊椎動物に向かって一直線に進化したように書かれている。しかし,実際には動物や植物は,それぞれ特定の動物門(phylum)をめざして進化した訳ではない。
もうひとつは,節足動物と脊椎動物への進化の主軸からそれぞれのグループ(phylum)が分かれたような図になっているが,実際にはそれぞれのグループの「線」は真上に伸ばさねばならない。節足動物,脊椎動物と横並びの図を描かなければならない。
しかしながら,図 18 を見ると、各動物門の系統関係(phylogenetic relationship)がわかりやすい。また,動物の進化が大きく 2 つの方向に分かれていることもよくわかる。2 つの方向に分かれて進化しただろうという根拠は,胚発生(embryonic development)における細胞分裂のパターン(らせん卵割と等分割)や,結果としてふ化する幼生(larva)の形態の違い(ベリジャー型幼生とディプリュールラ型幼生)が存在することにある。
図 18 の左側の経路で,刺胞動物から扁形動物が進化したことはほぼ間違いないだろうが,扁形動物から環形動物までの進化についてはよくわかっていない。大事なことは,先カンブリア時代から古生代にかけて絶滅した動物群が多いため,進化の経路がぼやけていることだ。
脊椎動物の方向も,刺胞動物から棘皮動物までの過程で,地質時代のいつ何が起きたのか,謎は深まるばかりである。
節足動物の基礎になった環形動物も先カンブリア紀に誕生したのだろう。新しい環境への適応に成功すると,あとはその適応様式をずっと維持し続けるのが基本的な進化のパターンであるが,環境は時間がたつと大なり小なり変化する。変化にうまくついて行けた動物は,形態も変わる可能性がある。逆について行けなかった動物は絶滅するのかもしれない。環境変化が大きければ,絶滅する動物も多いと考えられる。
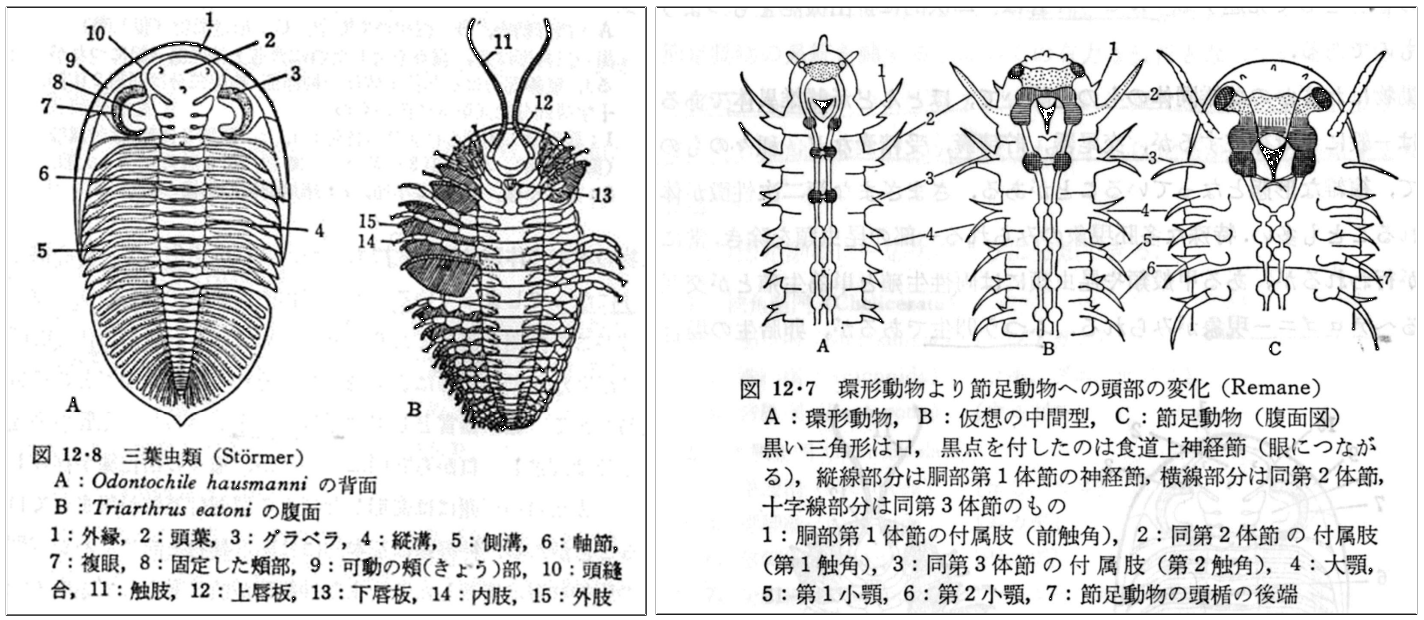
図 19(左)三葉虫(もっとも古いか不明だが,地球上に出現した初期の節足動物)。(山田・西田・丸山(1982)進化系統学。裳華房)
カンブリア紀から二畳紀の化石として産出。体は多数の体節(body segment)から成る。先端の 4 つか 5 つの体節が癒合して頭部を形成しているが,内側(腹側)の付属肢はそのままの形で残っている。体節構造が生まれた理由としては,捕食を免れるために体の装甲(甲羅)が厚くなったことが考えられる。三葉虫は古生代には繫栄しただろうが,中生代に入るころには絶滅している。こんなものを食べる捕食者はいなかっただろうから,絶滅の原因は,生息場所の大きな環境変化の可能性が高い。
図 20(右)環形動物から節足動物への予想される進化(形態から見た仮説)。(山田・西田・丸山(1982)進化系統学。裳華房)
節足動物の起源が、ゴカイのような環形動物にあったかは不明であるが,私は Remane の仮説(図 20)がわかりやすくていいと思う。ゴカイのような環形動物の先端 3 体節が癒合し,(上から見ると)頭部になった。一方,各体節の付属肢はそのまま残り,第 1~第 3 顎脚として捕食に機能している。十脚甲殻類では,各体節にある二叉型の付属肢は,体の前方から顎脚,歩脚,遊泳脚,尾扇の 4 つのタイプに変化している。
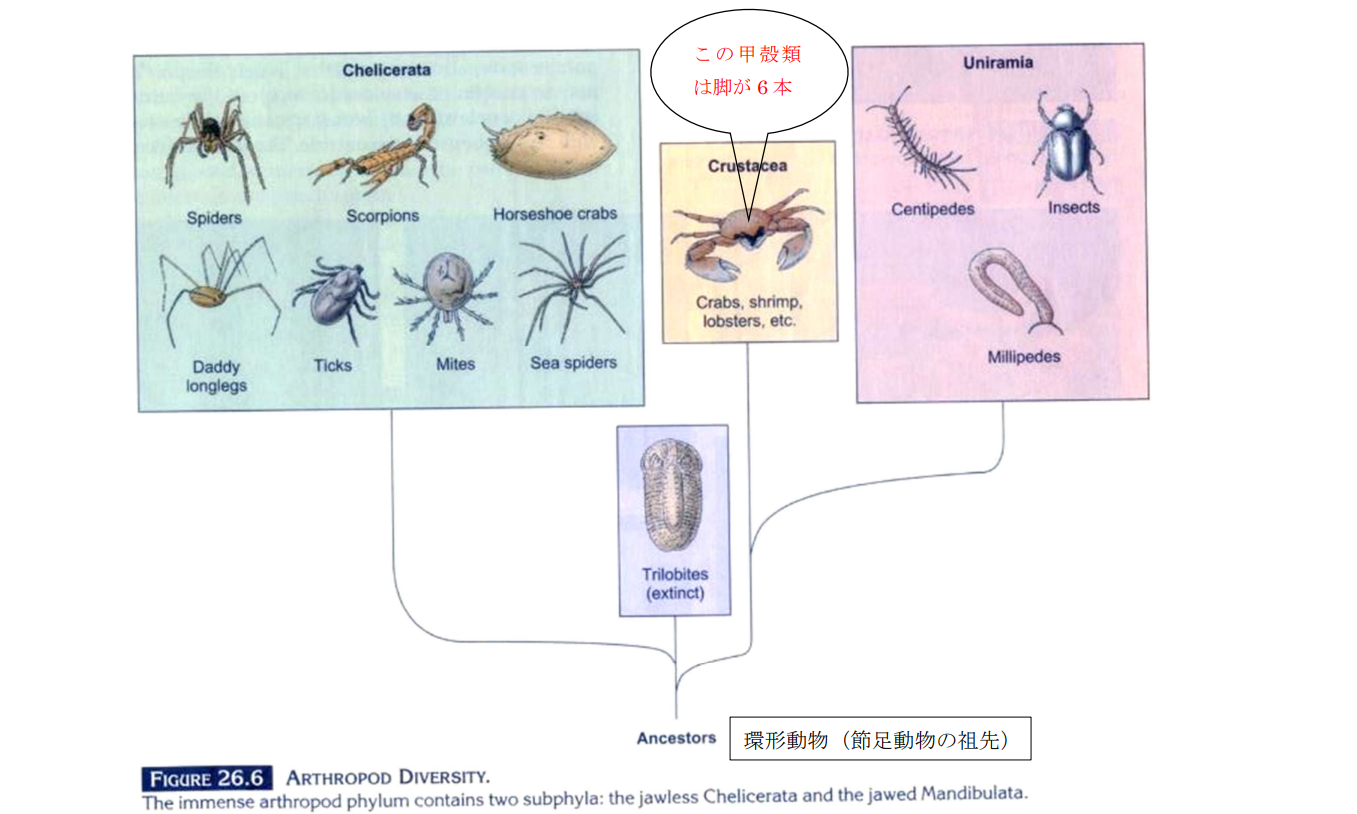
図 21.節足動物の多様性。三葉虫を含めて節足動物(Arthropoda)は,古生代に環形動物の仲間(種類は不明)から進化した可能性が高い。節足動物の進化の方向は 2 つに大別される。ひとつは鋏角亜門(Chelicerata)であり,カブトガニが含まれる。もうひとつは大あご亜門(Mandiburata)であり,十脚甲殻類(十脚目)は甲殻綱の中に含まれるが,この教科書では昆虫の含まれるUniramia から独立して扱われている。(図 21 の出典:Ferl, R.J., and R.A. Wallace (1996) Biology. Harper Collins College Publishers)。この教科書にはいたるところに間違いがあるが,独創性の高い本にはよくあることである。絵を描く人(著者とは別)が,カニの脚は昆虫と同じく 6 本だと思ったのだろう。この本は(日本の)中学,高等学校の教科書に強い影響を与えている。教科書には,間違った図がそのまま出てしまっているところがある。転載するときには注意が必要である。ただ,日本の社会は権威主義が横行しているので,間違いが指摘されてもうまく対応できないと思う。対応する人の選考が悪いということ。
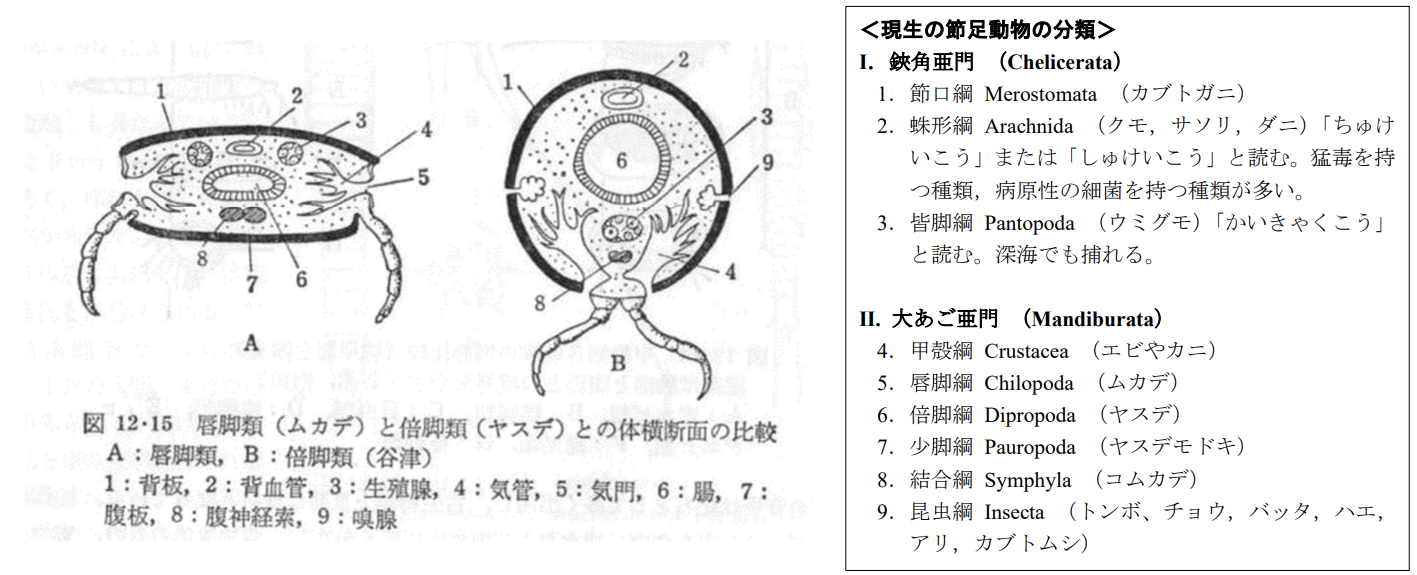
図 22.唇脚綱(ムカデ) と倍脚綱(ヤスデ)の付属肢の生え方の比較(山田・西田・丸山(1982)進化系統学。裳華房)
唇脚綱の動物(ムカデ)と倍脚綱(ヤスデ)の動物の付属肢(歩脚)の生え方には,大きな違いがある。ムカデ(唇脚綱)の体表(外皮)は,多少厚くなったクチクラが体の上半分(背版)と腹面(腹板)に分離し,その間から付属肢が生えている。一方,ヤスデ(倍脚綱)では,付属肢の基部(何という名前の節かは知らない)は左右でくっついていて,その間に腹板は見られない。
ムカデの場合には,各体節の側面から付属肢が出ることにともない,左右の付属肢が離れるため,その間のつなぎとして腹板が形成されるのだろう。体側から付属が出ることにより,各体節は上下に平たくなり,左右の付属肢の間隔(interval)が相対的に長くなるだろう。左右の付属脚の間隔が長くなれば,地面に付属肢の先をつけたときに体全体が安定する結果をもたらしたのではないだろうか?体が安定すれば,それだけ地上を歩行する速さが増すように思える。
一方,ヤスデの場合には,左右の付属肢にかかる体重を合体した「根本」で支えることになり,体全体はムカデに比べてやや不安定な支え方になるだろう。左右の歩脚のバランスが悪ければ,その分歩く速さは遅くなるかもしれない。実際に歩行する速度は,ヤスデに比べ,ムカデの方が圧倒的に早い。
唇脚綱と倍脚綱が地質時代のいつ頃に別れたかは定かではないが,歩脚の出方が,唇脚綱と倍脚綱で大きく異なることは,グループ(綱)を分けるための重要な指標と考えてよいのではないだろうか?十脚甲殻類の歩脚や顎脚にもグループによって大きな違いがみられる。

図 23.ロブスターの第 3 顎脚,第 1 歩脚(鋏脚),第 2 歩脚。付属肢の基節は左右でくっついている。基節は胸部に沿って前方を向いていることに注意。
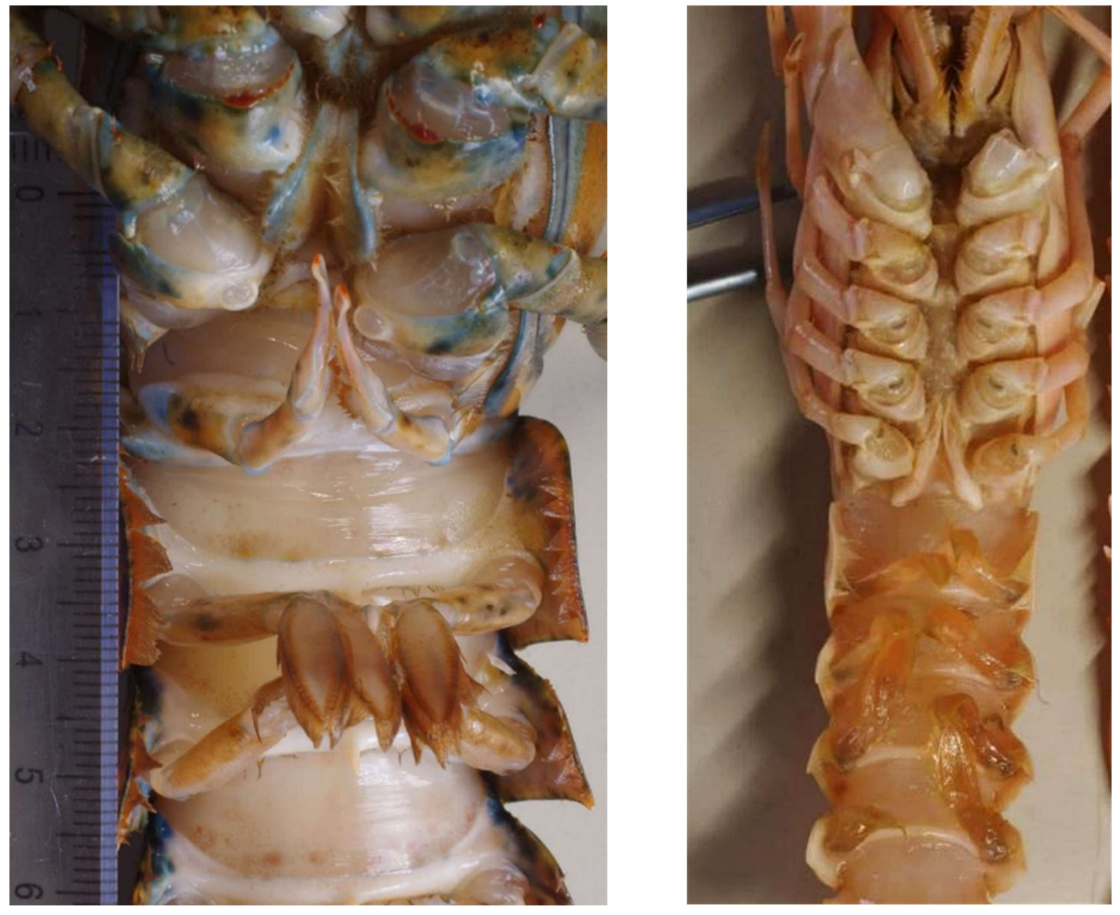
図 24.(左)ロブスターの第 4 歩脚と第 5 歩脚,遊泳脚。(右)アカザエビ(オランダ産)の第 3 顎脚,第 1~5 歩脚,および第 1 から第 4 腹肢。互いにつき方はよく似ている。どちらも腹板(図 22)は発達していない。

図 25.アカザエビの歩脚と遊泳肢。左は韓国産,右は土佐湾産。韓国産のアカザエビには,明瞭な胸部腹面プレート(唇脚綱でいう腹板)がみられる。オーストラリア産と韓国・日本産のアカザエビには,腹板の発達に違いがあるように思える。

図 26.サメハダヘイケガニの胸部腹面。腹板の面積が拡大し,各歩脚の基節は横(外側)を向いている。歩脚の出る位置は,唇脚綱(図 22)とよく似ている。す早い歩行に適した脚の構造になっている。カニ類の胸部腹面を見ると,カニ類はロブスターの祖先から進化したのではないことがわかる。