
1.Introduction
自然科学における研究力と教育力は,教科書に書かれている知識をできる限り多く吸収すれば,それで十分に身につくという訳ではない。例えば,大学入試センター試験を基礎にして人間を序列化する社会で,個人が良い位置を占めるには,知識を多く詰め込むことが有利に働く。しかし,研究現場や教育現場をよく見れば分かるように,人は知識を応用する力と創造力がなければ,結局は自分をごまかしながら組織を運営して行かざるを得ない。
うまく自分をごまかせた者は,社会から尊敬のまなざしで見られる。一方,うまくごまかせなかった者は,他人から揶揄される。うまく自分をごまかせなかった者は,自己の性格が歪み,結果として陰湿ないじめを発症する人も少なくない。私の周囲には,そんな人たちが多かったのかもしれない。そこまで人間歪むのかと考え込むほど,人格の崩壊した人たちをたくさん見てきた。(かく言う,自分も危ない時期が何度かあった。)自分の都合しか考えない人が混じると,職場の人間関係が急速に悪化するのだろう。
知識を応用する能力はどのようにしたら育つのだろうか?一流のパイロットといわれる人たちを見てみよう。一流のパイロットは,旺盛な知識の習得意欲に加え,多様な実戦的訓練(practice)を積み重ねている。戦前の海軍兵学校などは,まさにその典型であろう。野球にしてもサッカーにしても,強豪校は難度の高い実践的訓練を積み重ねている。現代の受験戦争も,知識の習得という本来の目標を逸脱し,受験に勝利するための厳しい実践的訓練を重ねている。平和な世の中になった証拠なのだろう。
強くなるためには,知識の習得に加えて,目的に合った実践的訓練は不可欠である。多様な実践的訓練の中には,体育会系的訓練というのもある。体育会系的訓練は,それで能力を伸ばせる者は良い。しかし,私のように失敗が多い者にとっては,失敗する度に罰が課せられる。だから組織に対する忠誠心は低下する。過剰なルール,過剰な指導,そして意味不明な精神主義に従えるほど器用ではない。
研究者や教員も同じようなところがある。センター試験や大学の授業の成績が良かったからと言って,すぐに実践で使える人材になる訳ではない。厳しい訓練に耐えてこそ,一流になれるのは間違いない。一方,厳しい訓練をさぼれば,いかに基礎学力があろうとも,やはり自分をごまかしながら生きる道が待っている。
厳しい訓練は,大学や多くの教育現場では,敬遠される傾向が益々強くなっている。些細な失敗でも訴えられ,たちまち処分やクビが議論される世の中である。恐ろしくてやっていられない。現在は,大学や教育現場で実施される実戦的訓練の多くは形がい化しているように思える。幼稚な学生が増加している状況で,指導する側の身の保全を考えれば仕方のないことだと思う。
社会に出てから実践で使える人材に成長するためには,大学や大学院に在籍している間に,自分で自分を鍛えるのが一番良い。やり方はいろいろあるが,海軍兵学校や陸軍士官学校に在籍する訳ではないから,当然自己流になる。しかし,自己流であっても,自分がうまく鍛えられた人は優秀なパイロットになれる可能性が高まる。優秀な研究者としてその道を歩んでゆくこともできるだろう。
・・・で,ろくに能力もないお前(私のこと)は,大学や大学院の中で何を鍛えたのか?私の場合には,いわゆる訓練期間というのはなかった。直接実戦に参加することで「訓練」とした。研究題目は,指導教員からの指示を受けて決めたものではない。だから,勝手なやつだとか何とか,さんざん悪口は言われた。そんな悪口(陰口)の山に耐えるというのも訓練のひとつではあるが,私のやった最も強力な訓練は,研究成果を出版できる論文にする作業であった。私は,原著論文(original paper)の作成には膨大な時間を費やした。それでもまだ論文をうまくまとめ切れていないが・・・。
私の持っている興味・関心は,ムツゴロウさんとよく似ているかもしれない。しかし,ただ一つ大きく異なる点がある。ムツゴロウさんは動物とのふれあいを一般向けの本として出版しているのに対し,私の方は原著論文にこだわり続けている点である。私の書いた本が売れるなどというミラクルは,万にひとつも起きる可能性はない。だからムツゴロウさんと同じ道を歩むことはできなかった。もちろん後悔はしていない。
一方,原著論文ならば,努力次第で自分を主張する道は開けると思った。(お金は儲かりませんが・・・。)日本人があまり書かない「introductionかなり長め」の面倒くさい論文には,私の主張がいっぱい詰まっている。しかも,英語と日本語の単語(word)の対応に消費する時間も多い。そんなややこしい論文の作成をサポートできるAIが出現するのはまだ当分先になるだろう。
原著論文を書くことに加え,私は物の「原点」を探ることが好きだ。これも誰かに教えられた訳ではない。生物進化の原点として,中生代における節足動物,特に十脚甲殻類(decapod crustaceans)の進化を探ってみたい。
十脚甲殻類には,クルマエビ(prawn),イセエビ,ウチワエビ,多くのエビ類(shrimp),タラバガニ,ズワイガニ,ガザミ,ザリガニ(ヨーロッパではよく食べられる),ロブスターなどが含まれる。いずれも水産関係(農学部,水産学部,各地の水産研究所や博物館)では特に関心の高い動物群である。進化の原点を探る上で,節足動物(Arthropoda),特に十脚目甲殻類(Order Decapoda)の体の基幹構造(fundamental structure)の形成に焦点を当ててみたい。生物の構造(形態)には,機能における類似性(相似器官)と系統的な類似性(相同器官)が入り混じっている。そこに主観の入る余地が残る。
何だ!そんなことはHaeckel(反復説で有名な人)以来,もう150年間も研究されて,すべてがわかっているのではないか?文献だって山ほどある。そんな時代遅れの学問を蒸し返して何をやるつもりなのか?・・・いやいや,何百年研究されてきたからと言って,節足動物の進化のメカニズムには謎が多い。
今まで多くの研究者は,系統と進化の謎には目をつむってきた。生命の起源や生物の進化には,人知の及ばないことがあると・・・。一方で,時代が進めば新しい研究方法も導入される。節足動物の体の基幹構造については,付属肢発現に関する遺伝子解析が最近注目を浴びている。付属肢発現の遺伝子解析や分子系統学的解析の後押しを受けて,基幹構造の構築という観点から節足動物(十脚甲殻類)の進化を見直してみたい。私の目標は「進化」の解明にある。一方,今の自分は分析力が衰え,各所に浪花節的思考がはびこり始めている。付属肢発現の「分子生物学的メカニズム」を手掛けるのは無理だろう。遺伝子発現の機構は,進化を説明する「道具」として使いたい。
話は動物分類学(taxonomy)と動物系統学(phylogeny)に戻る。多くの方々は気づいていないかもしれないが,両者はまったく方向の違う学問であることを強調しておきたい。分類学は,生物界を主に形態(見かけ)に基づいて整理する学問であり,リンネ以来ずっと社会に定着している。特別な知識や技術がなくても,少し直感が発達していれば学問ができる。多くの図鑑類,NHKの自然紹介番組,そして牧野富太郎を見ればわかるように,一般の人たちの受けは非常に良い。だから分類学は,社会貢献としては見栄えがするのであるが.問題は分類の際に使われる「直感」である。生物の進化は収斂的なグループもあれば,拡散的なグループもある。直感でやると,大別する基準(形質)が,グループによって異なってしまい,結果として見事に自然に騙される。
たとえば,分類学者は,硬骨魚(フナ)と軟骨魚(サメ)を分けているが,ともに魚の仲間という認識だろう。だから直感としては,硬骨魚と軟骨魚をひとくくりにしてもよいと思っている人は多いだろう。しかし,両者はデボン紀に分かれ,以来全く遺伝子の交流はなく現在に至っている。遺伝的には大きな違いのある生物なのだが,魚という形は共通である。一方,硬骨魚の系統からは,デボン紀に両生類が,三畳紀にハ虫類が出ている。起源が古いにもかかわらず,何で軟骨魚と硬骨魚は見かけが似ているのか?逆に,起源が新しいにもかかわらず,何で硬骨魚,両生類,ホ乳類の形態はこんなに違うのか,生物世界のパラドックス(意図に反する結果)を考えてみたい。
2.撮影と執筆の基本情報
<撮影場所> 西表島(干立の浜とユツンの浜)。11月の大潮(2004年),夜間の干潮時(真夜中の0時から3時ぐらい)。
<撮影と記事の執筆> 三枝誠行(NPO法人,生物多様性研究・教育プロジェクト常任理事)
<撮影機材> 風景はPENTAX K-rにTAMRONの18-200mmズームレンズ(Aspherical XR DiⅡ)をつけて,接写はK-rにSIGMA 50mm Macroをつけて撮影したと思う。K-rは完全に壊れたので数年前に廃棄した。K-rは15年間ぐらい使用したと思う。
<Key words> 十脚甲殻類,基幹構造,付属肢,生物進化のパラドックス。
3.参考文献
・Brooks, H.K. et al. (1969) Treatise on Invertebrate Paleontology, Part R: Arthropoda 4. The Geological Society of America and the University of Kansas.
・Castro, P., and M.E. Huber (2005) Marine Biology (fifth edition). McGrow Hill Higher Education.
・Factor, J.A. (ed.) 1995. Biology of the Lobster Homarus americanus. Academic Press.
・Forest, J., and J.C. von Vaupel Klein (ed.) 2004. Treatise on Zoology – Anatomy, Taxonomy, Biology. Vol. 1: The Crustacea revised and updated from the Traité
de Zoologie. Brill.
・Futuyma, D.J. (1998) Evolutionary Biology. Third Edition. Sinauer Associates, Inc.
・Herrick, F.H. (1909) Natural History of the American Lobster. Bulletin of the Bureau of Fisheries.
・広島大学生物学会(編)1971. 日本動物解剖図説。森北出版。
・木村資生(1988)生物進化を考える。岩波新書。
・西村三郎(編)1995.原色検索日本海岸動物図鑑 [Ⅱ] 保育社。
・Saigusa, M., and K. Oishi (2000) Emergence rhythms of subtidal small invertebrates in the subtropical sea: nocturnal patterns and variety in the synchrony with tidal and lunar cycles. Zoological Science 17: 241-251.

図 1.根鰓亜目(Dendrobranchiata)のエビ(prawn)。橙色が歩脚(5 対),黄色が遊泳肢(5 対),それに尾扇が付属肢となる。受精卵はばら撒き型。

図 2.根鰓亜目のエビ。潮が上げてくると砂から体を現す。頭部にある長い凶悪なトゲが Dendrobranchiata の特徴。飛び跳ねると捕食者に刺さる。

図 3.根鰓亜目のエビ。体節の色や模様から見てクルマエビに近いか?潮が上げ始めると体が砂の上に出る(潮汐時計を持つ)。眼はビデオライトの反射でよく見える。懐中電灯だと,探すのに時間がかかる。歩脚はアキアミ(いつも泳いでいる十脚目)とよく似た構造と思う。
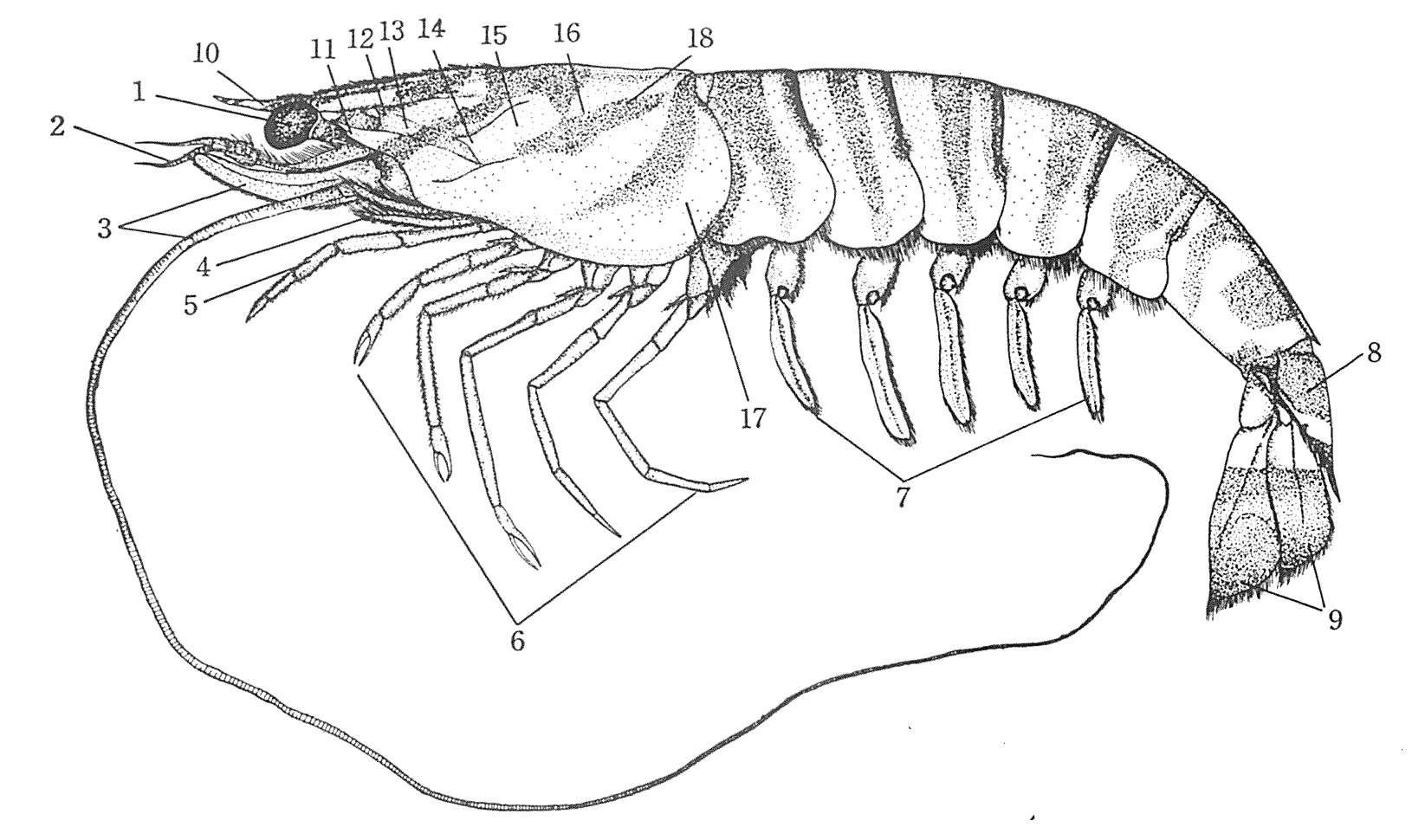
図 4.根鰓亜目(クルマエビ)の体の基幹構造。体は体節(body segment)で区分けされ,それぞれの大切に 1 対の付属肢(appendage)がつく。十脚甲殻類では,頭部から胸部にかけての 10 体節が融合して表面上は一つの体節のように見える。しかし,甲羅の内側では体節構造が残っており,第 1 触角(2),第 2 触角(3),第 1 顎脚(見えていない),第 2 顎脚(4),第 3 顎脚(5),第 1 歩脚から第 5 歩脚(6)が生えている。腹部の体節はそのまま残り,それぞれの体節からは 1 対の付属肢(遊泳肢)(7)が生えている。腹部第 6 節では,遊泳肢は退化して消失。尾部は,第 7 腹節(8)から尾扇(9)が生えている。頭部先端の凶悪な一本棘(とげ)が根鰓亜目の特徴。(日本動物解剖図説「クルマエビ」の項目を転載。)
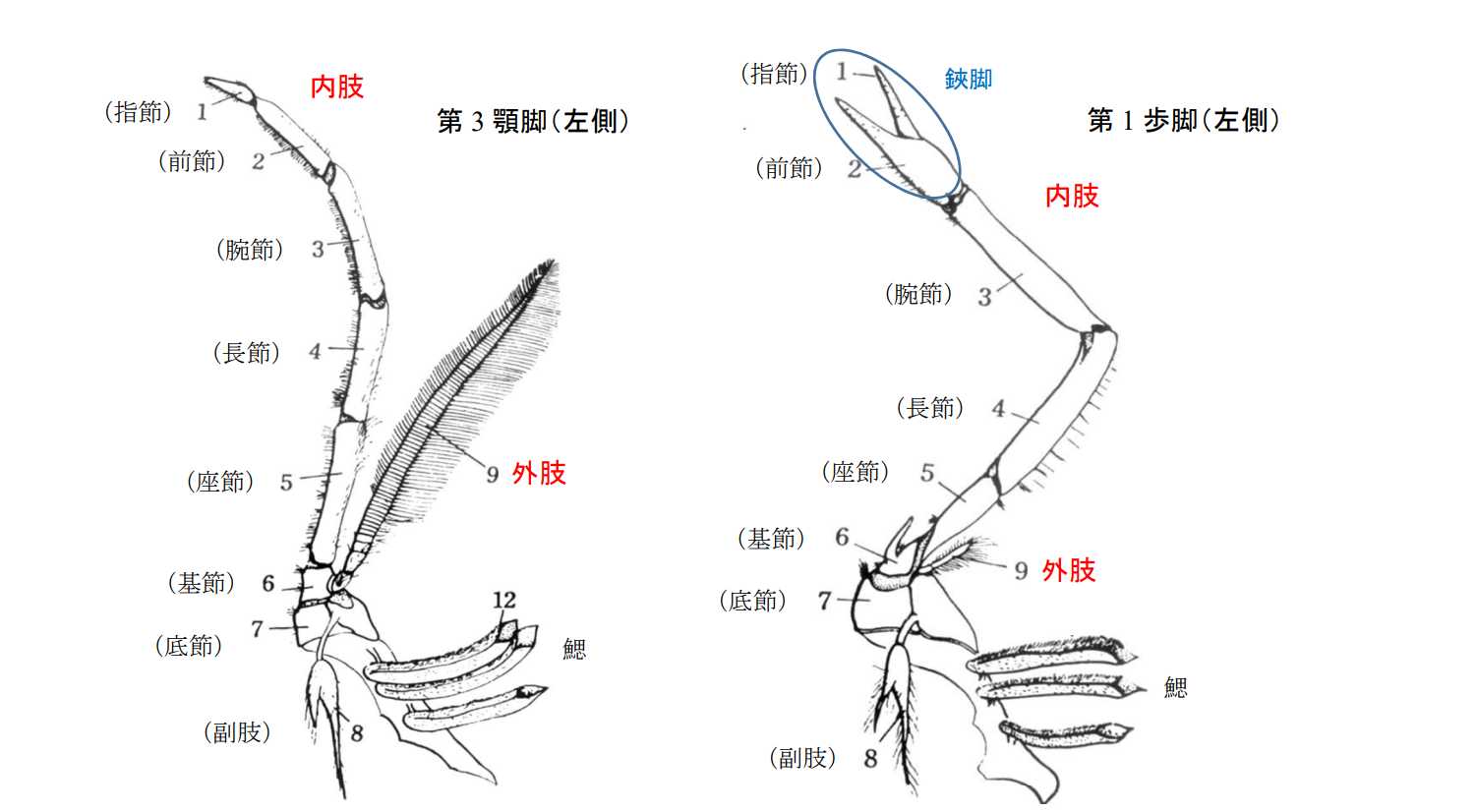
図 5.クルマエビ(根鰓亜目)の付属肢。(左)第 3 顎脚。(右)第 1 歩脚。(日本動物解剖図説「クルマエビ」の項目を転載)
十脚目の起源は,オキアミのようなアミ類だろう。二畳紀末の海産動物の大量絶滅(2 億 5 千万年前)の後,外洋で生き残ったオキアミが,超大陸パンゲアの東にあったテチス海の内湾で底生生活に入り,十脚目の中で一番原始的な根鰓亜目が誕生したのだろう。2 億 4 千万年前のできごとと予想している。
根鰓亜目の十脚類では,第 3・第 4・第 5 付属肢が顎脚(第 1・第 2・第 3 顎脚)に,第 6・7・8・9・10 番目の付属肢が歩脚(第 1・第 2・第 3・第4・第 5 歩脚)に分化(形態と機能の両方)している。しかしながら,顎脚と歩脚の構造は,形態の違いはあっても,基本的な構造(基節の部分で内肢と外肢に分かれる)は同一である。分類学者は形態の細かい違いばかりに注目するため,全体の大きな流れを見失っている。関心が無いようだ。

図 6.根鰓亜目のエビ(prawn)の生息場所。
採集場所は西表島のユツンの浜だったと思う。ユツンの浜は,島の北東側にある。ユツンの浜は,何も標識がないので,知らない人はどこに車を止めて,どこから入るかわからないと思う。車を道路沿いの空き地に止めて,道路沿いの入り口(すぐ近くに陥没口があるので注意)からジャングルの中を 10 分ほど歩けば浜につく。
小道には,ベンケイガニやオカヤドカリ(十脚目)がたくさん見られる。石の下にはサソリモドキがいる。雨の日にはサキシマハブも活動しているかもしれない。西表島はどこに行くにしても,長靴は必需品である。ただし,宿泊棟に置いてある長靴を使う場合には,事前に中をよくチェッ
クする必要がある。でっかいムカデが入っていることがある。
熱帯域や亜熱帯域に生息する動物は,夜行性(nocturnal)の種類が多い。西表島においても,夜間には昼間には見られなかった多くの生物を目にすることができる。
しかしながら,夜間に見られるといっても,潮が引かなければ,観察したり採集したりすることが難しい。潮の干満(ebb and flow of tides)のパターンは,1 年間隔で大きく変化する。潮の干満は,平均 12.4 時間周期で繰り返しているが,潮位(tidal amplitude)は,季節によって大きく異なる。
1 か月に 2 回ほどある大潮(spring tide)の期間には,満潮時の潮位は最大になり,かつ干潮時の潮位は最小になる。干潮時に関しては,春から夏にかけては日中の方が夜間に比べてよく潮が引く。一方,秋から冬にかけては,日中よりも夜間の方がずっとよく潮が引く。潮汐表を見ていただきたい。
西表島で海岸にすむ生物を観察しようと思えば,夜に潮がよく引く 10月から 1 月の大潮の時に海岸に出ればよい。ただし,ただし,西表島の夜間の海岸は大変怖い。海岸の前方には小浜島の光が見えるが,反対側は真っ暗なので,どちらの方向に陸があるのか全く分からなくなる。しかも,潮が上げてくると水の上をダツがバシバシ飛び跳ねている。

図 7.根鰓亜目のエビ(prawn)。図 1 と同じ種類か?潮が上げているとき(夜間の満潮の前後)は海にライトを当てると飛び跳ねるのがわかる。

図 8.根鰓亜目のエビ類(3 種類)。潮が上げてくると砂から体を現すので,ビデオライトがあれば簡単に採集できる。油でいためて食べられる。

図 9.こき使われてくたびれ果てる研究協力者の学生。11 月の大潮のころは,一番潮が引くのは午前 0 時から 3 時ごろになる。夜の 11 時時ごろ宿泊棟を出て 3 時間ほど採集し,それから帰るので,宿泊棟(琉球大学熱帯生物圏研究センター西表研究施設)につくのは朝になる。

図 10.抱卵亜目(Pleocyemata)のエビ(shrimp)。根鰓亜目のエビ(prawn)と違って体はずんぐりしている。受精卵は担卵毛に付着。

図 11.抱卵亜目のエビ(shrimp)。抱卵亜目のエビのもうひとつの大きな特徴は,砂に潜らずに底に積もった砂,転がっている石,砂地に生えている海藻(写真は褐藻類)の上を歩いたり泳いだりしている点である。根鰓亜目のエビとは,形はよく似ているが,全く別な動物である。

図 12.抱卵亜目のエビ。
夜間になるとサンゴ礁の砂地,サンゴ礫やサンゴ塊の上を歩いている。図 10 と図 11 のエビと同様に,根鰓亜目の頭部にあった凶悪なトゲは縮小し,代わりに触角(これも付属肢)の内肢が「へら状」になって前方に突き出ている種類が多い。
荒い砂に潜らなくなれば,歩く際に障害物にぶつかることは多いだろう。衝撃を避けるには,剣のような棘よりも,クッションになる「へら」や「ホウキ状の毛の束」の方が役立ったのだろう。
抱卵亜目のエビ類の起源は,根鰓亜目のエビと考えてよいだろう。根鰓亜目のエビが,多くは砂に潜るタイプだったのに対し,サンゴ塊の隙間に入って生活する種類が現れただろう。一方,スーパーで 1パック 200 円ぐらいで売っているガラエビがある。ガラエビは抱卵亜目だと思うが,頭部の棘(rostrum)がよく発達していて根鰓亜目のエビ類とは形がよく似ている。その他,見かけは根鰓亜目に似ているが,抱卵亜目に属するエビの種類は多い。
受精卵が担卵毛に付着するエビ(抱卵亜目)は,最初は 1 種類から始まったのか,数種類いたのかはまだわかっていない。根鰓亜目とガラエビタイプの基幹構造はよく似ている。抱卵亜目のエビ類の起源は,根鰓亜目のように思える。
中生代・三畳紀の前半,2 億 4 千万年ぐらい前に出現したのではなかろうか?
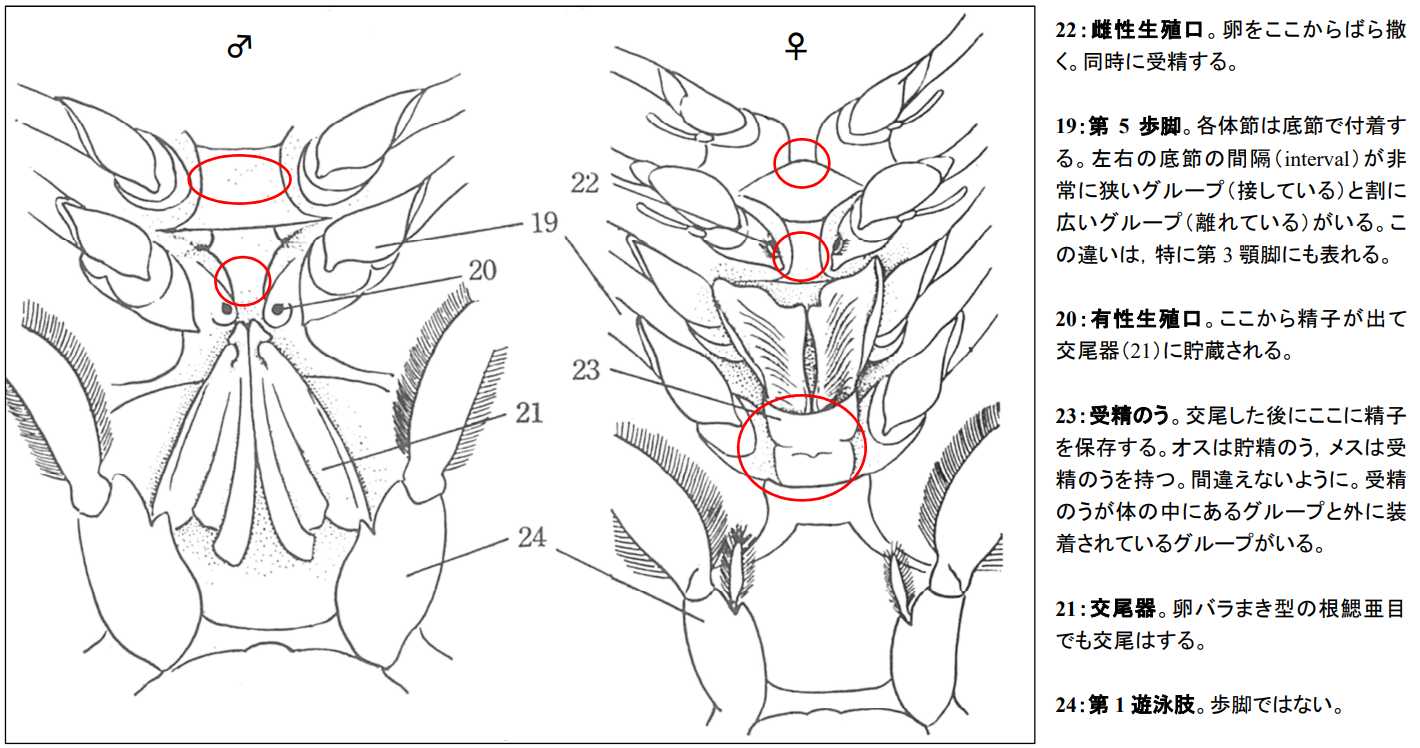
図 13.クルマエビ(根鰓亜目)胸部腹面のスケッチ。左がオスの胸部,右が雌の胸部を示す。(日本動物解剖図説「クルマエビ」の項目から転載)
十脚甲殻類の外部形態は,収斂(しゅうれん)が激しい。言い換えれば,体の基幹構造は,様々な環境に適応しても,大きくは変化しないことを示しているだろう。単なる想像であるが,基幹構造の発生(development)に関しては,遺伝子突然変異が修復されやすい機構があるのかもしれない。一方,形態形成(metamorphosis)が進んでから生じる遺伝子突然変異は,修復されにくいのかもしれない。形態形成の初期に起きる遺伝子突然変異は,体の構造を大きく変えてしまい,今までに少しずつ積み重ねてきた環境への適応を台無しにしてしまう可能性が高いような気がする。形態形成の後期ならば,形態のバラエティーが豊かになって,従って機能のバラエティーも豊かになって,さらに環境への適応も進む可能性がある。
一方では,基幹構造も進化とともに変化するだろう。例えば,歩脚の付け根(底節)の間隔は,グループによって左右で異なるかもしれない。

図 14.抱卵亜目のエビ。やはり体はずんぐりしている。受精卵は担卵毛に付着して胚発生が進む。腹部の先はくびれが強い。尾は根鰓亜目のエビとよく似ている。腹部から尾部にかけての形態は,アカザエビ,ロブスター,ザリガニとは大きく異なる。私はそれぞれの生物がどこから来たのか興味がある。

図 15.抱卵亜目のエビ。体はずんぐりして腹部は先端が大きくくびれている。イソギンチャクの触手の間に隠れている。抱卵亜目として祖先が地球上に現れてから(三畳紀中期)形態は大きくは変わっていないだろう。抱卵亜目のエビとしてひとつの完成形と言える。

図 16.県道からユツンの浜に出る道。こういう山道は子供を先頭にして歩かない方がよい。西表島に限らず,山の中はどこでもすぐ道に迷う。

図 17.ユツンの浜(大潮の干潮時)。八重山諸島のサンゴ礁原は,潮が引けば歩きやすいところが多い。こういうところなら子供たちを連れて行ける。ただし,親はいつも子供のそばを離れないように。写真の左側に赤い服を着た人が見える。手を引いてやればそこまでぐらいなら歩いて行けるが,それより先は子供連れでは危険が伴う。西表島は美しい島だが,昔から事故も多い。何をするにも親の判断が生死を分けることを覚えておいてほしい。