
1.Introduction
8月13日(日)は,昼から和中(吉備中央町)の基地の草刈りをした。草刈の直前に,林の中からカカカカカカ・・・・という幼鳥の低い鳴き声を聞いた。上空にはブッポウソウの姿が見えた。飛んでいたのはおそらく親で,幼鳥は基地の林に立ち寄って鳴き声を発していたのだろう。立ち寄った際に親にエサねだりをしていたのかもしれない。どこの巣箱で育った幼鳥か不明だが(B-01か?),わざわざ和中の谷に立ち寄って,私に渡り(migration)のあいさつに来たということではなさそうだ。しばらくして,親子ともども西の方角(宇甘川の方)に飛び去って行った。
吉備中央町では,今年(2023)巣立ちが一番遅かったのは,上田西の松田さん(R-20)の所だろう。R-20のペアは,200m北にあるσ-08の巣箱にいて産卵も行われた。しかし,σ-08を放棄して松田さんのところの巣箱で産卵を始めた。時期はかなり遅れて3つ卵が生まれた。松田さんから連絡をいただいて,8月6日に3匹とも巣立ったようだ。しばらくは池の周囲の林にいただろうが,もう旅だったかもしれない。また,池の東側にあるR-02のペアと争いを繰り広げたのかもしれない。R-02では,巣立った幼鳥は2匹ぐらいだったと思う。
近澤峰男さんにいただいた野鳥のファイルには,四季折々の野鳥の写真が収められている。8月は暑いこともあってか,撮影された野鳥の種類は少なかった。野鳥の中には,足根中足骨(ふしょ骨)が長い種類が多い。鳥類の足根中足骨は,見かけ上は人間(ホ乳類)の脛骨(足の脛の骨)に似ているが,素性は全く異なる。足首の先にある「足」の骨の一つが伸長してできている。だから,鳥にはヒトで言う「手のひら」とか「足の甲」に対応する部分がない。野鳥では,足根中足骨の先にあるのは,手の「指骨」であり足の「指骨」である。
では,なぜ野鳥では足根中足骨が発達したのか?ヒトも野鳥も同じ2足歩行である。違いは,ヒトはベタ足で歩くことが多いのに対し,野鳥の祖先である2足歩行性のハ虫類(エリマキトカゲに似た鳥盤類?)は,つま先立ちで走ったと推察される。2足歩行でつま先立ち歩きが,鳥への進化の中で足の骨の形態変化を起こさせた可能性がある。一方,前足は翼に変わったが,大腿骨と脛骨に関しては両生類やハ虫類とよく似た形態が維持されている。
2.撮影に関する基本情報
<撮影者と所属> 三枝誠行・近澤峰男(生物多様性研究・教育プロジェクト)
<撮影場所> 古座一枚岩(和歌山県東牟婁郡古座川町),
<Key words> 足根中足骨(ふしょ骨),二足歩行,ベタ足,つま先立ち,手(翼),足(後ろ足),足指骨,Archosauer。
<撮影機材> EOS 7D Mark II(Canon)with 600mm Lens (Canon)と思う。EOS 7Dの前に使っていたカメラ(機種不明)かも知れない。
<参考文献>
Futuyma D.J. 1998. Evolutionary Biology, Third Edition. Sinauer Associates, Massachusetts.
広島大学生物学会(編)1971. 日本動物解剖図説,森北出版

図 1. ケリ。羽毛に覆われている部分は脛骨。大腿骨は腹部に埋め込まれている。足根中足骨は長い。来住(兵庫県小野市)にて。平成 27 年 8 月 21 日。
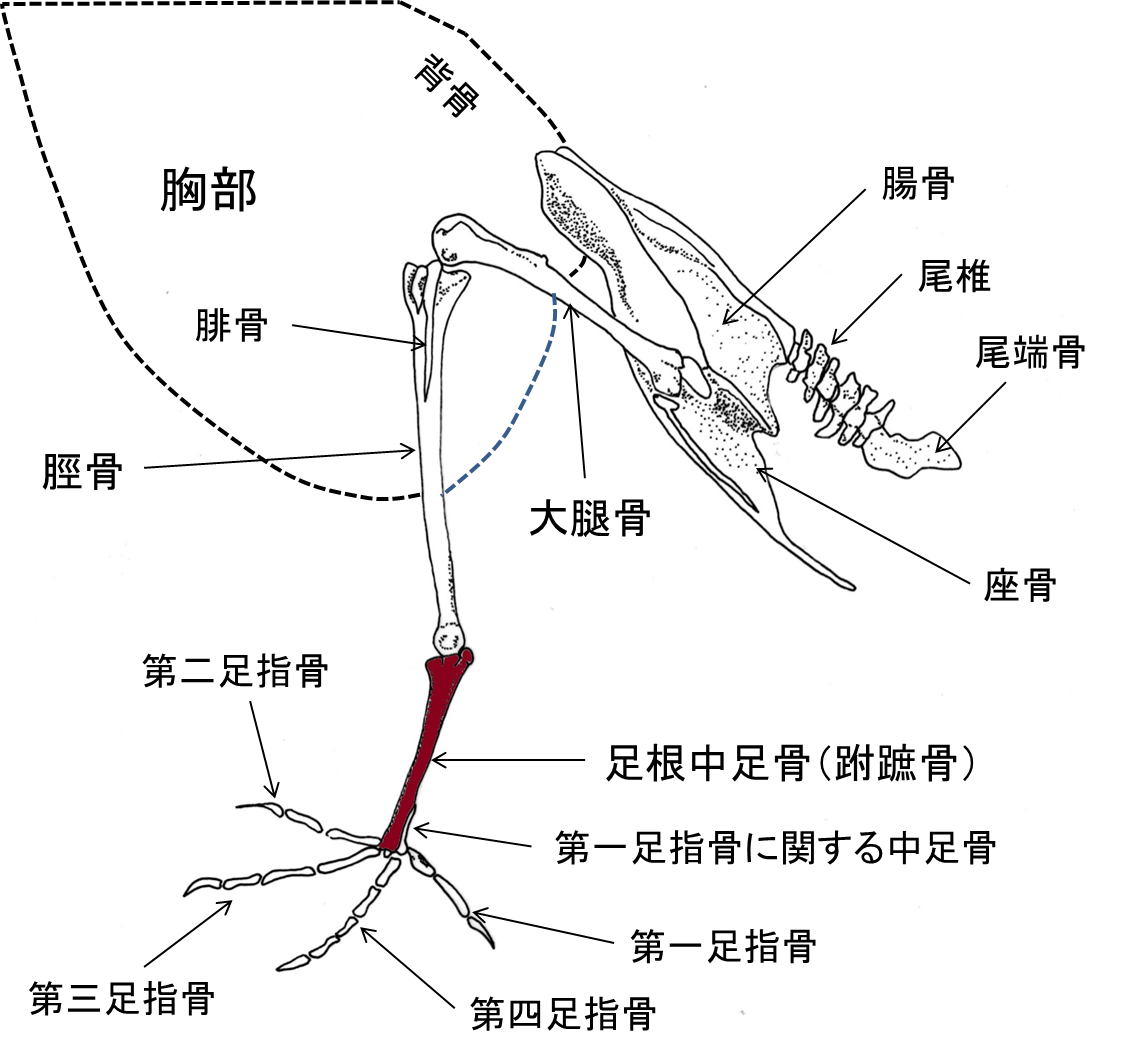
図 2. 野鳥(ドバト)の骨格構造。足根中足骨(茶色に塗った骨)はホ乳類で言えば足の甲にあたる。足根中足骨の先は第 1 足指骨から第4足指骨。
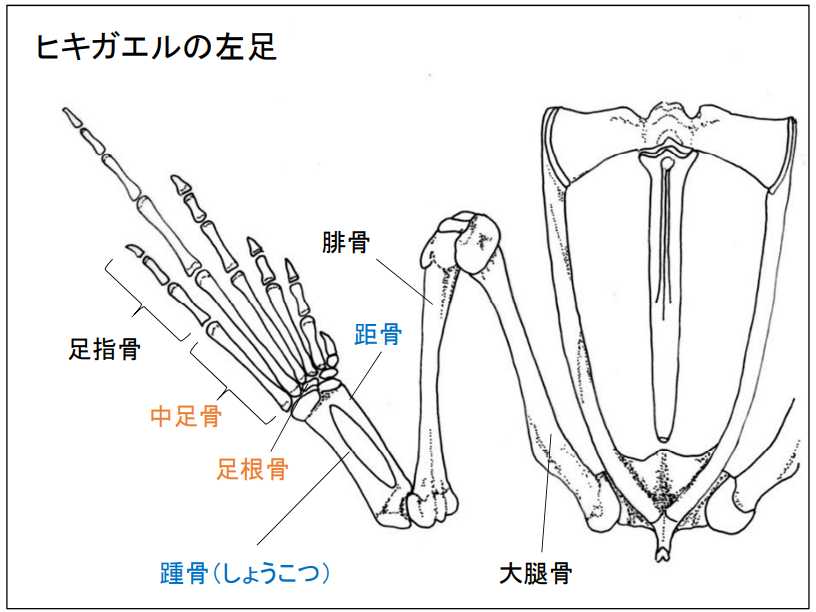
図 3. 両生類(ヒキガエル)の足の骨(左足)。
カエルやヒキガエルの仲間は,両生綱の無尾目(Anura)に分類される。尻尾はオタマジャクシの時に生えているのみで,オタマジャクシからカエルに変態する際に消失する。
無尾目の両生類の特徴は体のいろいろなところに現れ,後ろ足では足根骨と中足骨は,複数の骨で構成されている(図 3)。
面白いのは,腓骨と足根骨が踵骨(しょうこつ)と距骨(きょこつ)という 2 種類の骨を介してつながっていることだ。踵骨と距骨の形状は,イモリ(有尾目)やホ乳類の脛骨(けいこつ)や腓骨(ひこつ)と構造が似ているが,別物である。おそらく足根骨(図4)が変化したものだろう。ハ虫類は,両生類から進化したとされるが,カエルやヒキガエルのタイプの構造を引きついだグループは出現しなかったのだろうか?竜盤目の恐竜には,踵骨大腿骨 と距骨はなかったのだろうか?
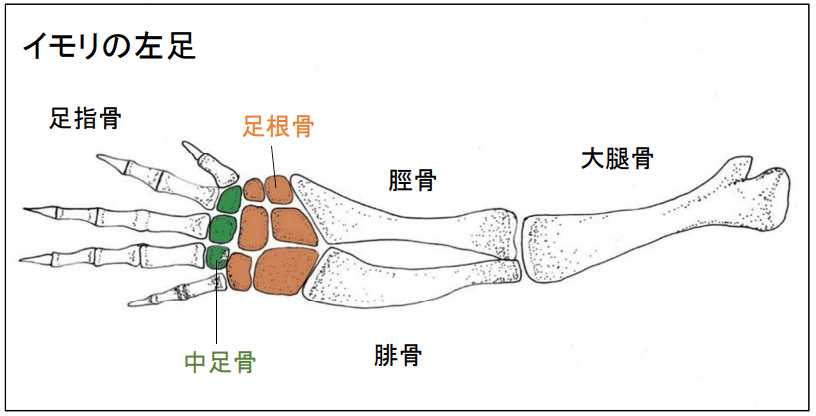
図 4.両生類(イモリ)の足の骨(左足)。
カエルの足とは,大きく異なっている。イモリの足の構造はホ乳類と非常によく似ている。言い換えれば,鳥類の足の構造とも大きく異なっている。イモリの足は,ホ乳類(シロネズミ)とよく似ているので,足の骨構造だけから見れば,イモリに近いグループからハ虫類を経てホ乳類に進化したグループがいた可能性は高い。ハ虫類は徐々にホ乳類に進化したようだ(いわゆるホ乳類型ハ虫類)。疑問は,ハ虫類を長くやってホ乳類に変ったグループと,ハ虫類の時期が比較的短かったグループがいた,なんてことはないだろうか。
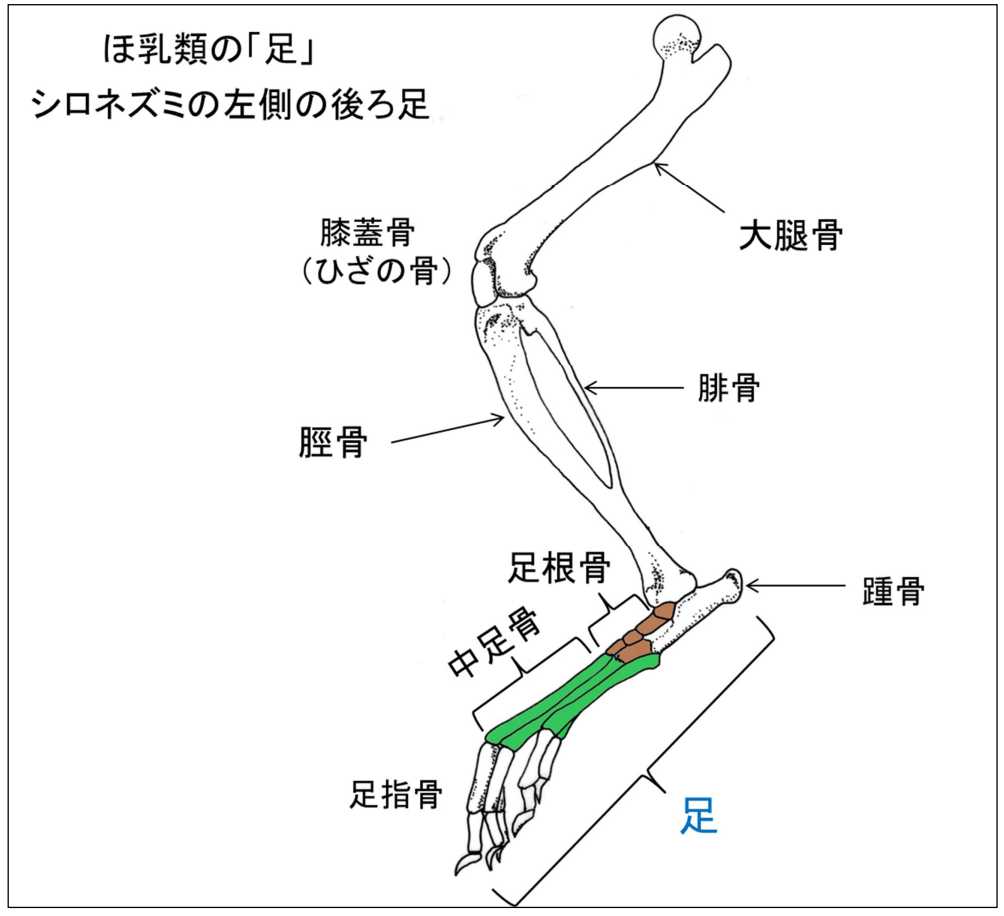
図 5.ホ乳類(シロネズミ)の足。
シロネズミの足の構造は,基本的にヒトの足の構造とよく似ている。
足の構造だけから見ると,イモリ型両生類の足は,カメの足と構造が似ている。トカゲやワニともよく似ているような気がする。
足の構造からみると,ホ乳類はイモリ型両生類から,トカゲ型のハ虫類を経由して現在の姿に至っている感じがする。
では,鳥類(Aves)はどこから進化したのだろうか?ワニが 2 足歩行をやったのは見たことがない。体が重すぎて,後ろ足ではとても自身の体を支えることができないだろう。キシノウエトカゲなども無理だろう。
そうなると,体の軽い鳥盤目のトカゲ(見かけからだと,キノボリトカゲやエリマキトカゲなどを連想するとよい。)から進化したとする可能性が浮上する。
2 足歩行のできるトカゲ類(恐竜に近いか不明)が,速く走るため,またできるだけ遠くを見るためにつま先立ちを繰り返した結果,足根骨と中足骨が一本になり,長さもぐんと伸びたのかもしれない。
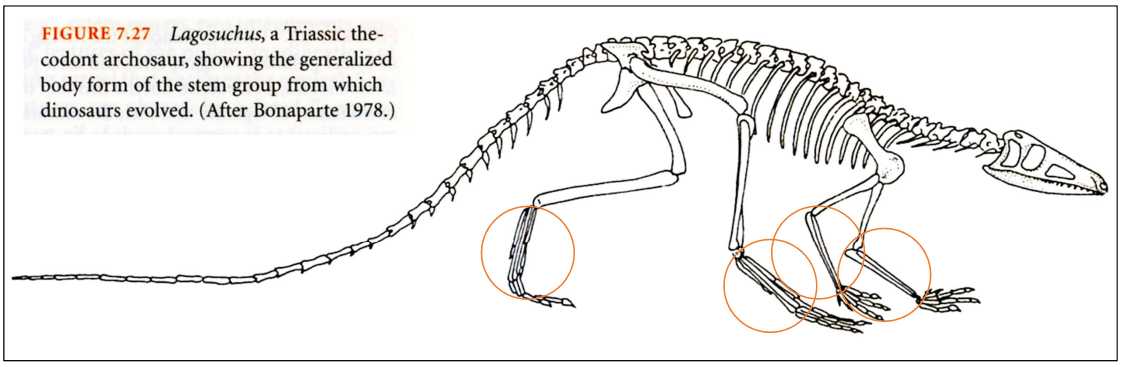
図 6.三畳紀の古恐竜の体の構造(Futuyma 1998 より転写)。
図 6 は,三畳紀に存在した古恐竜(Archo は「古い」という意味)の骨格構造を示している。手や足の骨の名称が記述されていないので,それらが両生類やホ乳類のどの骨に対応するかよくわからない。大腿骨と腓骨(あるいは脛骨)は,イモリ型両生類やホ乳類と対応するように思われるが,橙色の丸で囲った部分はどんな骨でできているかよくわからない。足根骨や中足骨が伸長している状態とみてよいのだろうか?あるいは,カエルのように踵骨(しょうこつ)と距骨(図3)が伸びているのだろうか?
鳥に進化したグループ(考え方のバイアスを避けるため,あえて調べていない。)では,すでにこの段階(archosauer)から体重が軽くなり,足根骨や中足骨(まだ複数の骨からできている)が伸長し始めていることが予想されるのだが・・・。
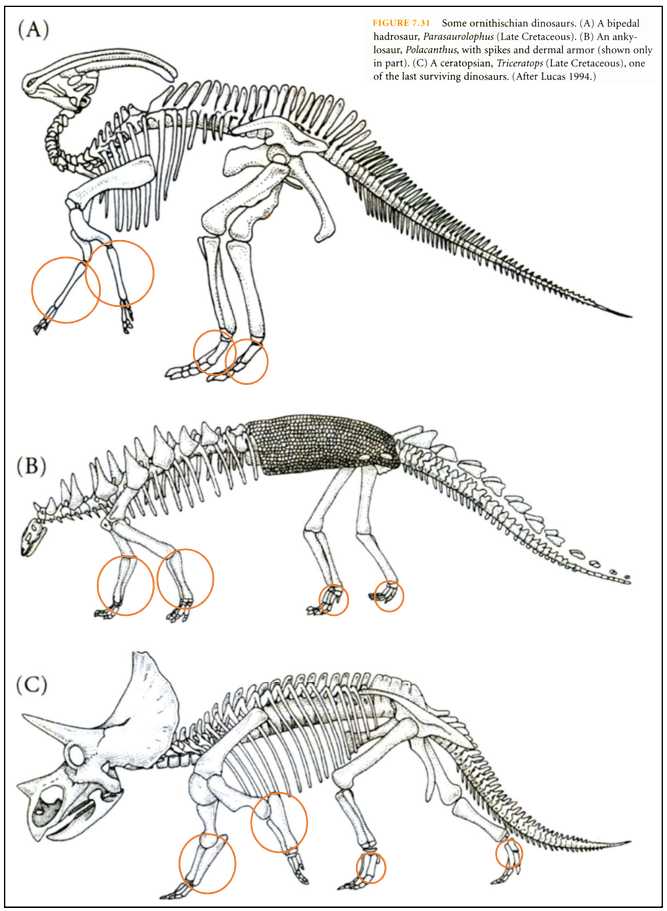
図 7.鳥盤目恐竜の足(Futuyma 1998 より転写)。
Ornithischian dinosaurs は,鳥盤目恐竜のことを言う。Cのトリケラトプスはよく耳にする名前である。トリケラトプスを含めて,バイペダル・ハドロザウル,アンキロザウルにしても,頭骨・背骨・尾骨は好き勝手に伸び放題になっている感じが面白い。
恐竜類は,骨盤の特徴から大きく「鳥盤類」と「竜盤類」の 2 つのグループに分類されているようである。鳥盤類は,骨盤の上部の骨(恥骨)が長く後下方に伸び,下部の骨(坐骨)に対し平行的に伸びている。現在の鳥の骨盤と共通した特徴を持っている。
一方,竜盤類はトカゲの骨盤に形が似ているということで名付けられたようだ。
頭骨・背骨・尾骨は進化の過程で好き勝手に形を変えていったが,鳥盤や竜盤は原始的な恐竜からあまり形態を変えずに続いている。
見かけにこだわると,生物の分類は,「人為分類」の側面が強くなる。綱(class)とか目(order)の場合には,グループに共通した特徴を選べば,自然分類の感じが強く出てくる。
鳥盤目恐竜の足(橙色の丸)は,脛骨や腓骨(図 5)でできている可能性が高い。重い体重を支えるには肥大化した脛骨や腓骨が必要だったのだろう
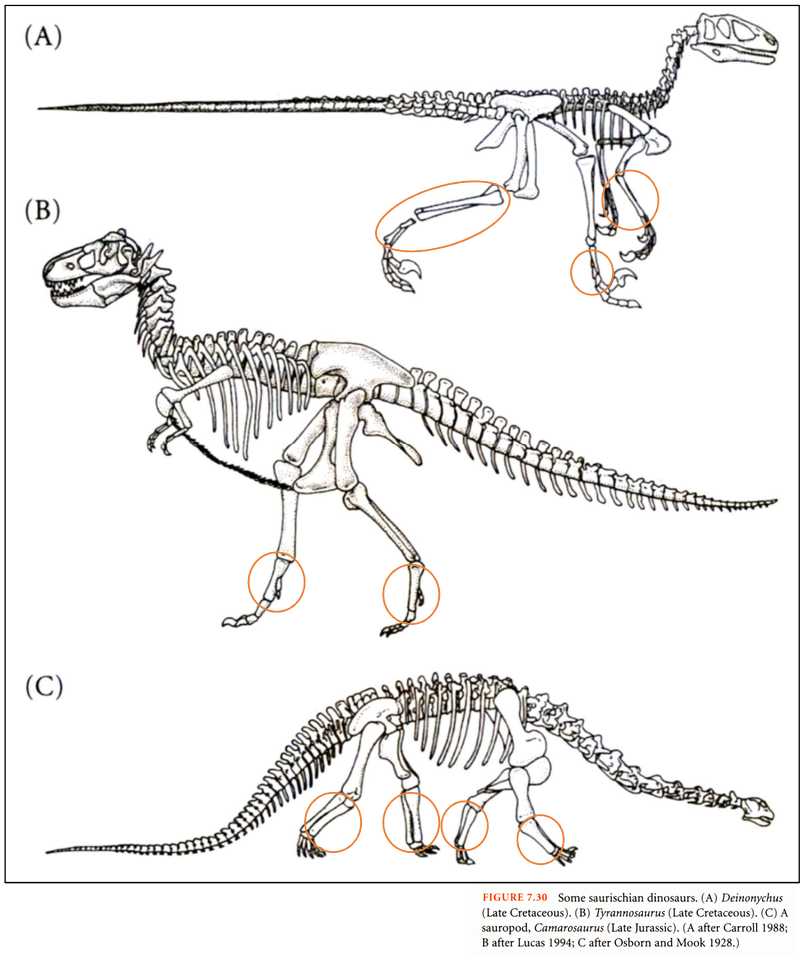
図 8.竜盤目の恐竜類(Futuyma 1998 より転写)。
白亜紀後期の竜盤目恐竜(A と B)は,2 足歩行だろう。C の恐竜はベタ足歩行と思われる。
2 足歩行(A と B)にしてもベタ足歩行(C)にしても,2 本または 4 本の足で重い体重を支えなければならない。足は,大腿骨,脛骨と腓骨,その下(橙色の丸)は足根骨と中足骨が合体して肥大化したか,あるいはどちらか一本になり太くなったか,どちらかであろう。2 足歩行(A と B)の場合には,力点(point of effort)は足根中足骨,支点(fulcrum)は足指骨の付け根,作用点(point of load)は足指骨の地面と接する部分になるのだろう。
特に足指骨の付け根の部分には強い負荷がかかるので,恐竜の足は痛風になっていたかもしれない。もっとも,ハ虫類がどれだけ痛覚が発達していたか不明であるが・・・。長い首,大きな頭部(脳は小さい)を支えなくてはならず,肩こりも相当なものだったような気がする。体が大型化したり,凶暴な捕食者に変身すれば,種(species)としては成功するのかもしれないが,個別の生物個体としては,体のいたるところに大変な負担を強いられることになると思う。だから,環境が変化すれば,短時間であっけなく絶滅ということもあるだろう。
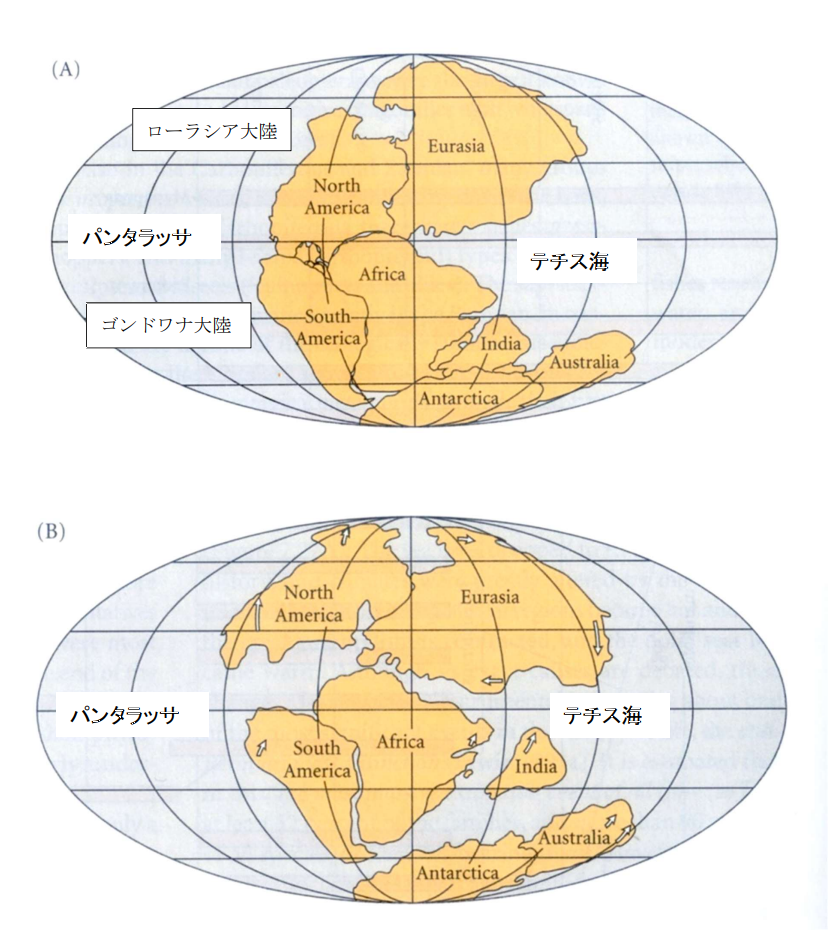
図9.三畳紀からジュラ紀にかけての大陸の分裂と移動(Futuyma 1998 より転写)。
A は三畳紀(Triassic),B はジュラ紀(Jurassic)後半の大陸を示す。なお,大陸の分裂と移動の詳細な年代に関しては教科書によって大きく異なる。
地球上には,古生代には一つの超大陸(パンゲア)が存在したと考えられている。パンゲアの東側にはテチス海が,西側にはパンタラッサ海が広がっていただろう。コリオリの力によって生じる海流は,北半球ではテチス海とパンタラッサの間を大きく右回り,南半球では大きく左回りに流れていたと思われる。
中生代(三畳紀)に入ると,パンゲアは大きく2つの大陸(ローラシア大陸とゴンドワナ大陸)に分裂したようだ。さらにジュラ紀に入ると,ローラシアはそのまま残ったが,ゴンドワナの方がインド亜大陸,アフリカ大陸,南アメリカ大陸,そして南極-オーストラリア大陸に別れた。
ジュラ紀には,テチス海とパンタラッサはまだ残っているが,特にテチス海周辺の海流は三畳紀とは大きく異なっただろう。
三畳紀からジュラ紀にかけてのパンゲアの分裂と大陸移動が,無脊椎動物(invertebrates)と脊椎動物(vertebrates)の両方に大きな変化をもたらしたのではないだろうか? この考え方を支持する多くの証拠が残っているはずだ。その証拠をできるだけ多く見つけたい。
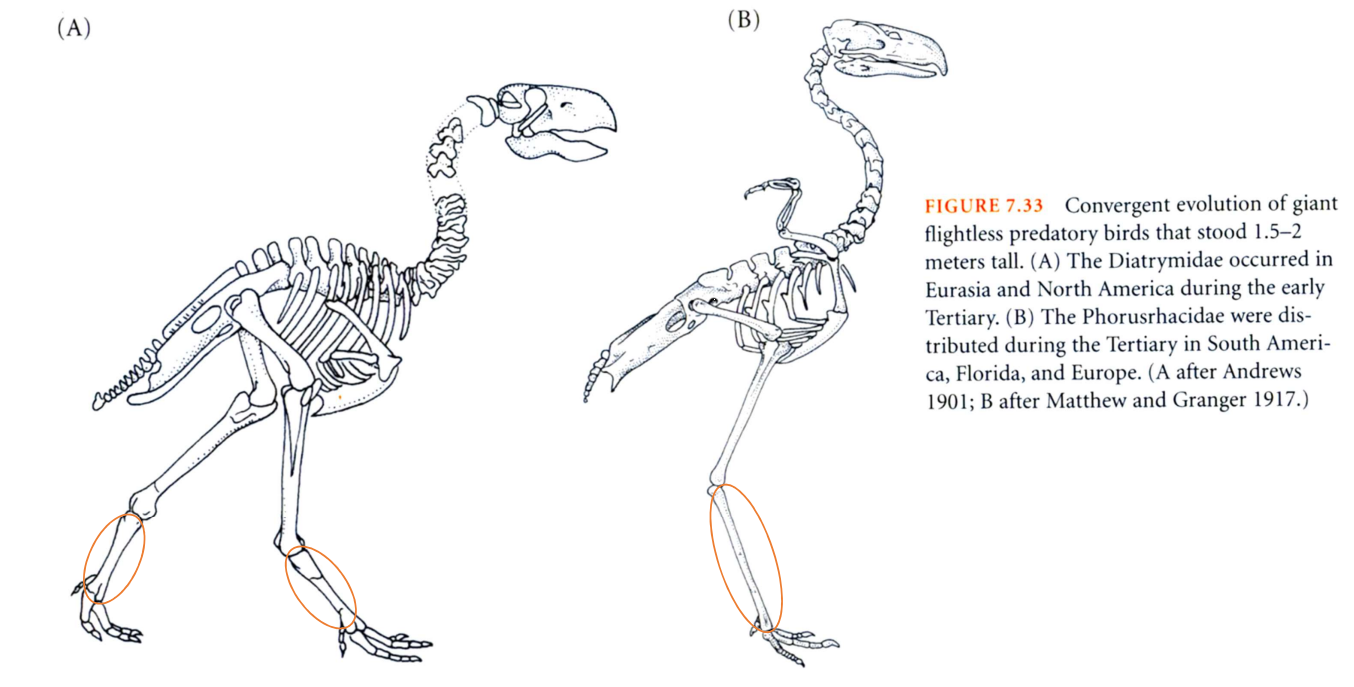
図 10.巨大捕食鳥類の収斂進化(Futuyma 1998 より転写)。
Diatrymidae(A)は三畳紀初期にローラシア大陸に分布した。horusrhacidae(B)の方は,三畳紀にゴンドワナ大陸の南アメリカからユーラシア大陸西端まで分布した。ともに飛ばずに陸上を走って移動するタイプの「鳥」であった。
鳥の出現はジュラ紀ではなかったかと思う。また,分離した大陸が現在の大陸の地名と対応できるのは,ジュラ紀も後半だろうから,図(FIGURE 7.33)の説明(figure caption)に三畳紀とあるのはジュラ紀の間違いだろう。
始祖鳥の中で翼はあるが,地上を走り回って獲物を見つけていた地上性の鳥(同じ種)が出現して,ローラシア大陸の方に進出したグループとゴンドワナ大陸の方に分布を広げたグループがいたのだろう。ジュラ紀の終わりごろに産出した化石を調べると,両者(A と B)は形態的に非常によく似ているということで収斂進化と記述されている。A も B も,橙色の楕円で囲ったのは足根中足骨であろう。ただ,B の方は現在の鳥類と似て洗練された骨格構造という感じを受けるが,A の方は原始的な特徴が強く残っている感じがする。収斂(しゅうれん)(convergence)と言うよりは,多様化(diversification)と言った方がよいのではないか

図 11.イソヒヨドリ。古座一枚岩(和歌山県東牟婁郡古座川町)にて。平成 27 年(2015)8 月 7 日。私はこの日は和中(吉備中央町)にいた。

図 12. 古座川の景観。古座一枚岩をインターネットで調べたら,こんなきれいなところでした。(https://www.wakayama-kanko.or.jp/spots/21/)

図 13.オオルリ。平成 22 年(2010)8 月 29 日,若杉原生林(岡山県西粟倉村大茅)で撮影。幼鳥か? 近澤さんは若杉原生林が好きだったみたいだ。

図 14.ソウシチョウ。平成 22 年(2010)8 月 29 日,若杉原生林にて。ソウシチョウは平成 17 年に特定外来生物に指定されたとのこと。

図 15.ヒバリ。平成 27 年(2015)8 月 21 日,来住町(兵庫県小野市)で撮影。私が見たらスズメにしか見えない。

図 16.ヤブサメ。平成 22 年(2010)8 月 29 日,若杉原生林にて。ヤブサメの「シーシーシーシィィィ・・・」という鳴き声はよく聞くが,ご本尊がこんな姿をしているとは知りませんでした。それと,皆さまお分かりいただけるだろうか?現代の野鳥は,古代の野鳥に比べて著しく洗練された姿に進化している。2 億年かけて,野鳥は姿・形ともに完成品になっていることをお忘れなく・・・。完成した形態なのだから,きれいなのは当然か・・・。