
2026年5月20日(水)
東京高等師範学校は,貧乏人家庭の子息が入学する学校だったのか?
私は,小さいころから生物が大好きであった。夏休みには,夕方になるとごみ溜めから大きなミミズを掘り,ウナギ針に通し,近くの小川に仕掛けた。朝になると親に起こされて,小川を見て回った。10個しかければ時々1~2匹はかかっていたと思う。体長30㎝もあろうかいう大きなアマゴ(伊豆半島に分布するのはヤマメではなくアマゴと思う。)がかかっていたこともあった。
朝食の前には,山の道沿いのクヌギの樹液を見てクワガタムシ(主にミヤマクワガタとノコギリクワガタ)を採集した。バケツに半分ほど採集できることもあった。樹液にはオオスズメバチも来ており,回避してクワガタを採集するのは大変に時間がかかった。朝食をとるとすぐに,向かい側の家の広い庭と自宅の周囲でアブラゼミを採集した。8月も15日(旧盆)を過ぎるとアブラゼミに代わってツクツクボウシが採集できた。クマゼミはほとんど見られなかった。昼過ぎると,天気がよければカワトンボやシオカラトンボを採集した。しかしオニヤンマの採集は子供には難しかった。盆を過ぎるとアカトンボの採集が多くなった。
朝食の前には家から500mほど離れた集会所でラジオ体操が行われた。これも行くのは大変嫌だったが,欠席すると怒られるので,いやいやながら手足を動かしていた。
昼過ぎると近くの川に泳ぎに行ったが,水は冷たく3分も入っていられなかった。特に私は,ツベルクリン検査でいつも擬陽性になったので,毎年肩にあの忌々しいBCG接種をされた。BCG接種の後はかさぶたができ,水に入ると接種した部分の表皮に直径5~6mmほどの穴ができ,中には真皮まで見えていた。冷たい水に入ると,言葉では言い尽くせないほど肩は激しく痛んだ。私の家族は,私が小学校を卒業するのに合わせて東京に出た。
中学校や高等学校(三流高)では成績は悪くはなかったが,ダントツと言えるほど良くもなかった。勉強の仕方がわからなかったことが原因だったと思う。加えて勉強することが自分の将来にとってどれほどの意味を持つか,よく理解していなかった。親が田舎(静岡県田方郡中伊豆町上大見)を離れなければ,私は大嫌いな農業を引き継ぎ,長い間つまらない人生を歩んでいたことは間違いない。「この虫キチガイじじい」とか言われて,通りがかりの子供らに小石を投げつけられることもあったに違いない。親が田舎を出なかったら,私が親の代わりに東京に出たと思う。もちろん好きな生物学の研究などとてもできはしなかっただろう。

図1.茗荷谷のキャンパスにある小さな池。今でも昔そのままに残されている。何という名前か知らない。タナゴを採集して遊んだ。青いイトトンボもいたような気がする。 初夏にはゴマダラチョウが飛んでいた。親の収入が一番少ない大学,つまり貧乏家庭の子息が入学する大学と言われていたが,私が入学するころには多少違ってきたようだ。入学試験のコツさえつかめれば,合格するのはそんなに難しい大学ではなかった。偏差値の高い大学をめざすのも良いが,重箱の隅をつつくような大学入試共通テストの対策に明け暮れたのでは,大学に入学後の勉強がおろそかになるだろう。今の日本の社会は,韓国や中国と同様に,大学に入ること自体が人生の目的になっている。一浪しても二浪してもいいから,入学するのが難しくない大学を選ぶという手もある。大事なことは,大学に入学してから仲間に迎合して遊んでしまわず,自分でコツコツと将来の勉強を進めることである。今はAIが発達しているので,うまく使えば大学での授業よりもずっと効率的に学習できる。
東京高等師範学校の入学試験問題(生物学)
つい最近,我が家の本の整理をしているときに,関口晃一著「いつの間にやら米寿を越えて」(アサヒ出版)を見つけた。もう10年ほど前に著者から我が家に送られてきたと思う。本をめくってみると,著者の少年時代から現在までのことが時系列で記されていた。少し気になったので読んでみると,何と関口晃一氏の人生は私と瓜二つではないかと思えてきた。関口晃一氏は,1919年に埼玉県北埼玉郡騎西町牛重で生まれ,東京高等師範学校(1952年廃校)と東京文理科大学(1953年廃校)を経て,東京教育大学と筑波大学の教員として教壇に立ち続けてきた人である。
関口晃一氏が1938年(昭和13年)に受験した東京高等師範学校(大学ではなく,旧制高等学校と同格)の入学試験では,生物学に関して以下の5つの設問があったと書かれている。試験時間はおそらく3時間で,受験者には試験問題作成委員(誰だったかは不明)が指定したサイズのわら半紙が5枚配られたであろう。印刷は刑務所で行われていると聞いた。具体的には,
・牛の反芻胃について述べよ。
・細胞の大きさについて記せ。
・植物の蒸散を調べる実験方法を記せ。
・水辺の生き物について君が観察したことを記せ。
・他に一題(詳細は不明)
今の人たちは「正解」に慣れ過ぎてしまっているので,こんな正解もない問題を出したと言って怒るかもしれない。
しかしながら,この入学試験問題は簡単に解ける。大事なことは知識の量を問うのではなく,論理的な整合性を問うための出題であることを理解することである。私はこの類の問題だと,9割は取れない(採点者の好みが異なるため,採点にばらつきが出る)にしても8割は確実に取れると思う。
こういう問題であれば,私はいくらでも書ける。昔は答案用紙の裏側まで書いてもよかったと思うが,裏側まで書くとボロが出て減点の可能性が高くなるので,表側だけでまとめる方がよい。
生物が85点,英語が70点,数学が50~60点,国語,理科(物理・化学・地学からひとつ)と社会が,それぞれ60~70点程度取れれば,大学入試共通テスト(当時はなかったが・・・)の成績が平均70点でも,個別試験で十分ばん回できる。私も東京高師ならば十分に合格できたと思う。今はどうかわからないが,かつての早稲田大学教育学部の入学試験問題(生物学)も,こんな感じではなかったかと思う。面白いことに,生物の成績が良いと他の試験科目の成績も良くなるのは不思議である。
東京高師の入学試験問題を見ると,確かにこの大学には貧乏人の子息が好んで受験してもおかしくなさそうな感じがする。泥臭い感じの試験問題である。物理や化学も似た設問だったろう。東京大学や京都大学であればもっと洗練された設問になるだろう。
個別試験は大学によって内容が異なるので,いわゆる「大学ランキング」にこだわらずに点数が稼げそうな大学を選ぶと良い。そういう大学だと自分が興味を持つ研究ができる可能性が高い。嫌いなことをうるさく言われながら研究するのは骨が折れる。

図2.1971年に乗船した「ひめゆり丸」。(許可を得て「時刻表ギャラリー」から転載。)「ひめゆり丸」は「おとひめ丸」より少し小さく,2,500トンぐらいの貨客船だったと思う。那覇港から乗船したのは3月10日の昼過ぎと記憶している。おとひめ丸の写真はたくさんあったが,ひめゆり丸の写真は探しても出てこなかった。私は那覇‒石垣間では船酔いに悩まされることはなく,昼夜関わらず船内をくまなく探索した。私と同じように,船内を徘徊する(?)若者はたくさんいた。南の島に行くという期待が,室内でじっと待っていられなかったのだと思う。私たちは石垣港で下りたが,この船は台湾の基隆(Keelung)まで行ったと思う。東京‒那覇間と同様に那覇‒石垣間も多くの若者が乗っていたが,台湾まで行った者は少なかったかも知れない。船は若い時しか乗るチャンスがない。大学生の時に西表島まで船旅をしたのは本当に良い経験になった。おとひめ丸とかひめゆり丸のような船に乗って,高さ5mもあろうかという大きなうねりにもまれ,激しい船酔いを体験する中で,旧式の貨客船に乗せられ南方(ソロモン諸島やニューギニア)に送られた兵士たちの無念に思いを馳せることもできる。辺野古に行かなくても,いろいろな平和学習が可能である。先日の事故では,平和学習を利用して高校生を反戦活動に参加させた疑いがある。これはまずいだろう。

図3.上空から見たフェリー(2024年7月3日)。石垣島から宮古島に向かうJAL便から撮影した。石垣か宮古のどちらの方向に向かっているか不明。今(2026)はフェリーで宮古島や石垣島に旅行する人は少ないと思う。長期間滞在するのであれば自家用車といっしょに乗船できるが,当然ながら運賃は高い。

図4.宮古島のサンゴ礁原(reef flat)。宮古諸島や八重山諸島の島々は,サンゴ礁に囲まれている。海岸からサンゴ礁の縁(白波が経つところ)までは, 200mから400mはありそうである。宮古島が隆起してから写真にあるサンゴ礁原が形成されるまでは,そんなに年月がかかっておらず,2,000年から5,000年ほどではなかろうか?サンゴ礁原はさすがにカラー写真でないと構造がよくわからない。また,船からは真横から見ることになるため,サンゴ礁原の広さはよくわからない。

図5.海岸に生息する動物プランクトン。海岸に突き出た桟橋で夜に電気をつけると,電灯の下にはたくさんの動物プランクトンが集まってくる。このサンプルは岡山県瀬戸内町牛窓の海岸で採集されたと思う。大きな種類はアミ(甲殻類)とイワシの稚魚(硬骨魚類),それに茶色いヨコエビ(甲殻類)と橙色のハルパクチクス(甲殻である。その他,黄色いプランクトンはクマセア(甲殻類),一番小さいのはコペポーダ(ミジンコ)である。種類は手前の2匹のクマセアを除いて不明。
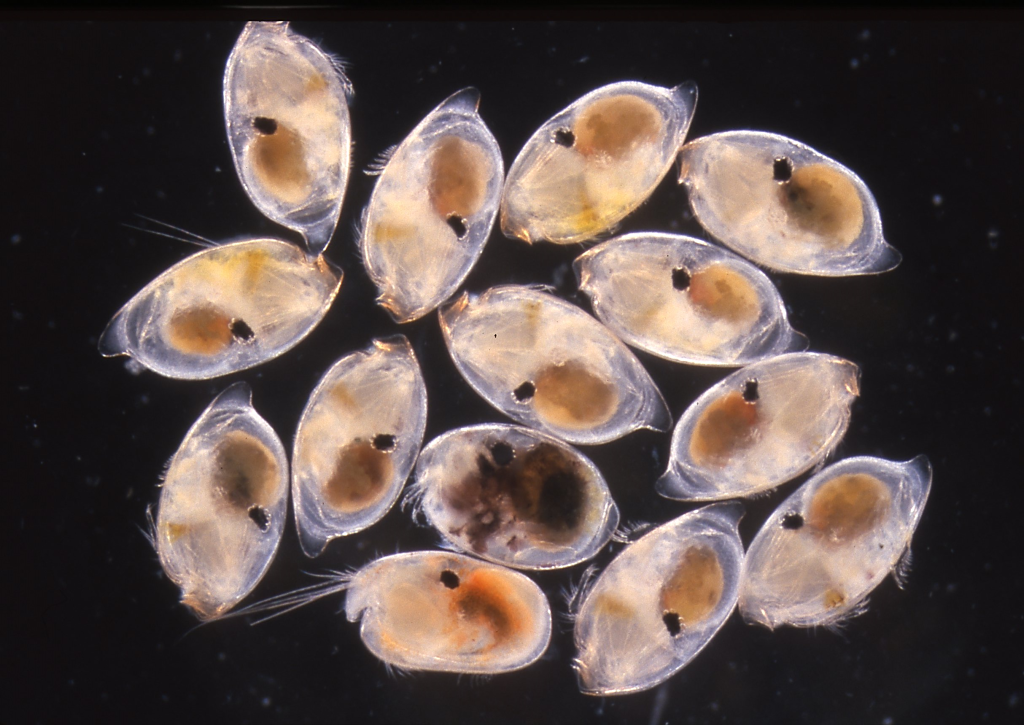
図6.動物プランクトン採集で捕れたウミホタル類。西表島祖納港で採集したと思う。ウミホタルの仲間は,甲殻類(Class Crustacea)の貝形虫亜綱(Subclass Ostracoda),ミオドコーパ目(Myodocopa)に分類される。3種類のウミホタルが混じっているが,この中で夜の海で光るのは,1種類だけ(中央の一番下の個体)。

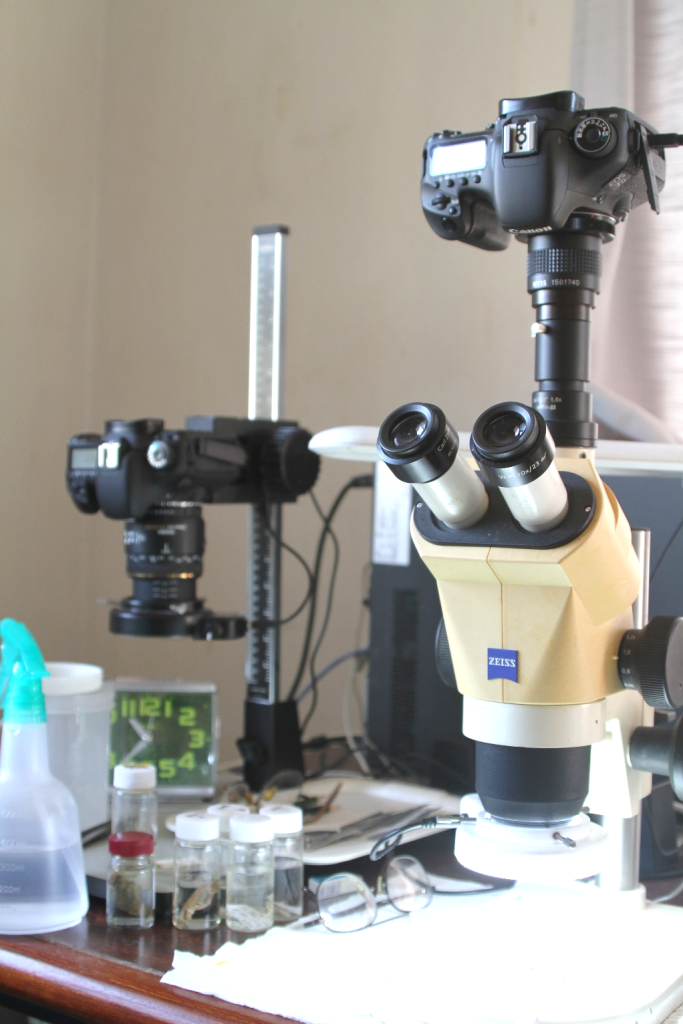
図7.動物プランクトンの撮影に使用される顕微鏡(左側の写真)。動物プランクトンの撮影は,暗視野照明の下で行われる。体の内部構造がある程度明確に写っていれば,種の分類も可能になる。左側の顕微鏡(双眼実体顕微鏡)の機種は,オリンパス SZX 12。レンズの汚れが激しかったので分解掃除をした。光学顕微鏡は,機種は古くても使い慣れれば十分にきれいな写真が撮れる。今はパソコンにつないで,モニターを見ながらカメラ(中古のCanon EOS 7D)のシャッターを切れる。ファインダーを除きながらシャッター・ボタンを押すのはものすごく大変である。顕微鏡システムは,業者に依頼せずに自分で工夫して作るのがよい。部品はインターネットで注文できる。自分で構築すると,長く使うことができる。右は落射光型の普通の双眼実体顕微鏡(Zeiss Stemi 2000)。カメラ(中古のCanon EOS7D)に接続するためのCマウント・アダプターがついていたので,撮影鏡筒をいくつか購入して自分でシステムを構築した。形態形成(morphogenesis)の研究には顕微鏡は必需品。

図8A.発光実験に使用したウミホタル(Vargula hilgendorfii)。
ウミホタルは灯火にも集まるが,大量に捕れる訳ではない。たくさん集めようと思えば,海岸にトラップをしかければよい。まずは近くの魚屋に行ってイワシでもサバでもよいから何匹か生魚を購入する。生魚は,骨ごと切り身にしてカニかごに入れ,海岸に投げておく。次の日に引き上げ,かごを上下に振ってやれば,下にある受け皿に大量の肉食性の甲殻類が落ちてくる。瀬戸内海だと,大量のウミホタルが採集できる。
他にもいろいろな種類の甲殻類が落ちてくる。左の写真には,大量のウミホタルが写っているが,右上には体長1 cmほどの肉食性甲殻類(スナホリムシ)が見える。スナホリムシは結構凶暴で,砂浜の海岸を歩いていると寄ってきて,ヒトの足の皮ふを食いちぎることがある。また,春に海藻の繁茂した内湾を泳ぐと,海から上がって体を見ると,お腹のまわりに大量のワレカラ(甲殻類)がくっついていることがある。そういう経験も楽しい臨海実習の思い出になる。
日本の大学の理学部・農学部・水産学部には,本学と離れて臨海実験所(呼び名は学部によって異なる)を持っていて,実験所に滞在して長期間海洋生物の研究ができる。昔は臨海実験所の利用者が多かったが,現在ではどこも閑古鳥が鳴いている。自然の生物を相手にして研究する人が減ったことがその原因である。

図8B. ウミホタルの発光。大量に集めたウミホタルを入れた水槽の両側に電極を置き,低電圧(10Vとか20Vとか・・・)をかけると,ウミホタルはいっせいに発光する。ウミホタルの発光のメカニズムは,分子機構のレベルで解析されている。今後は,解析の結果をどのように応用するか,そちらの方に研究が進むだろう。
苦労はするが,自分で新しい研究を始めることが一番就職のチャンスにつながる気がする。ウミホタルの発光のメカニズムなど,研究している人は少ないだろう。しかし,小さな分野だと,大事にしてもらえる。小さい分野だから,研究関係に就職のチャンスは少ないかと言うと,逆である。大きな流れ(例えば遺伝子発現の分子生物学的研究)に従えば,義理人情の人事が待っている。権威者のご機嫌を損ねて嫌われでもしたら,一発でドロップアウトになる。その意味では,臨海実験所に長期間滞在し,一人でコツコツと生物学の研究を行ってきたという実績は,高く評価される可能性がある。
くれぐれも「鶏口となるも牛後となるなかれ!」 と強調しておきたい。パーマネント・ポジションを得るまでは,ひたすら我慢して耐えるべし。我慢できなければ惨めな将来が待っている。アドバイスと称して「ウソ」をつく奴らとか,悪いうわさをすぐに広げようとする奴らがいる。そのうちに氏名を公表しても良いと思うが,そういう連中に限って公表された途端に被害者面をして訴えてくるので,無視しておくのが得策かもしれない。
図9A.臨海実習で講義を行う安井金也氏(広島大学向島臨海実験所)。
ウミホタルは,種名(Vargula hilgendorfii)とミオドコーパ目(Myodocopa)というグループの名称を指す場合の両方に使われる。ミオドコーパ目には何種類のウミホタルが記載されているか知らないが,どうしてVargula hilgendorfiiだけが光るかは謎のままである。
生成AIによれば,ウミホタルではルシュフェリンとルシュフェラーゼが海水中の酸素と反応した際に,高いエネルギー状態の分子ができる。高エネルギー状態が安定な状態にもどるときに,余ったエネルギーが青い光として放出されるとのこと。ホタルの発光も似たようなメカニズムで起きるが,発光は細胞内で起きるようだ。私の関心は,ルシュフェリンとルシュフェラーゼが海水中の酸素と反応した時に,なぜ高いエネルギー状態の分子ができるのかということである。ルシュフェリンとルシュフェラーゼがそれぞれにエネルギーを持っていて,それらが酸素を媒介として化学合成が行われた際に余剰のエネルギーが放出されるということなのだろうか?
もしそういうことであるならば,最初の生命(細胞)も同じようなメカニズムによってできた可能性がある。カギはペプタイドの自然合成にありそうだ。

図9B.臨海実習で海洋生物の講義をするRushan M. Sabirov氏(カザニ大学)。広島大学向島(むかいしま)臨海実験所にて。
日本の社会は,視野の狭い人が非常に多い。視野が広ければ,それだけ多くの客観的情報をつかむことができ,正確な判断ができる可能性が高まると思う。視野の広い人間になるためには,例えば教養教育科目の広範な履修が役に立つだろう。教養教育科目の特徴は,多少の知識をもとに「考える」ことをエンカレッジする学問であることだ。つまり,個人としての人間が将来どのように生きたらよいのかを考えさせてくれる学問と言えると思う。大学に入学して最初の2年間は,教養教育科目の履修に専念すべきだ(とまでは言っていないが)という文部科学省の方針には,私は賛成する。ただ,教養の生物学とか教養の物理学みたいな教科書は面白くない。「数学精義 Ⅰ・Ⅱ・Ⅲ」は大学で教養教育科目として優れた教科書だと思う。
安井氏もサビロフ氏も視野が広い。だから私のような武骨な人間でも相手にしてくれたのだと思う。サビロフ氏は,昨年(2025)お亡くなりになった。安井氏はまだ元気だと思う。安井氏と交流があるというだけで,向島臨海実験所は出入り禁止となってしまった。ビックリ・ポンの組織である。

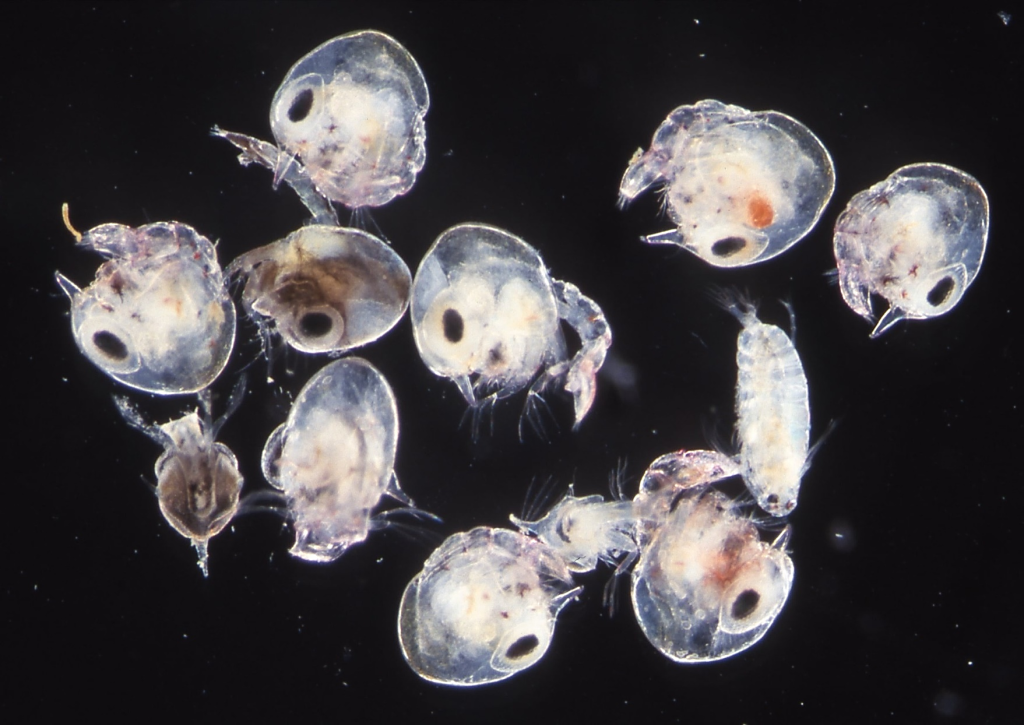

図10.灯火に集まる動物プランクトン。西表島の祖納港や美田良(みたら)の河口で採集した。左の写真は,イカ(種名は不明)の赤ちゃん。大きさ1 cm弱。右の写真は夜間の満潮時にメス親から放出されたカニ類のゾエア幼生(zoea larvae)。シオマネキ(ヒメシオマネキ,ハクセンシオマネキ,ベニシオマネキ) のゾエアだろう。小さい焦げ茶色のゾエアは種類不明だが,コメツキガニのゾエアの可能性がある。プレゾエアと言っている人たちがいるが,形態形成(morphogenesis)から見ればそんな段階(stage)はない。ゾエアでよい。大きなコペポーダ(体長500 µmほど)も混じっている。熱帯や亜熱帯の島々の森や海岸には発光生物が多い。西表島では,3月にヤエヤマヒメボタルの集団が,20時から30分ほどの間にいっせいに発光する。体内時計(サーカディアン・クロック))がある。イカは光る代わりに色素胞(収縮胞と呼んだ方がよい?)を持つ。ゾエアは発光もしない代わりに方向の決まらない泳ぎする。南の海には生物進化のメカニズムを研究するのに適した生物が多い。
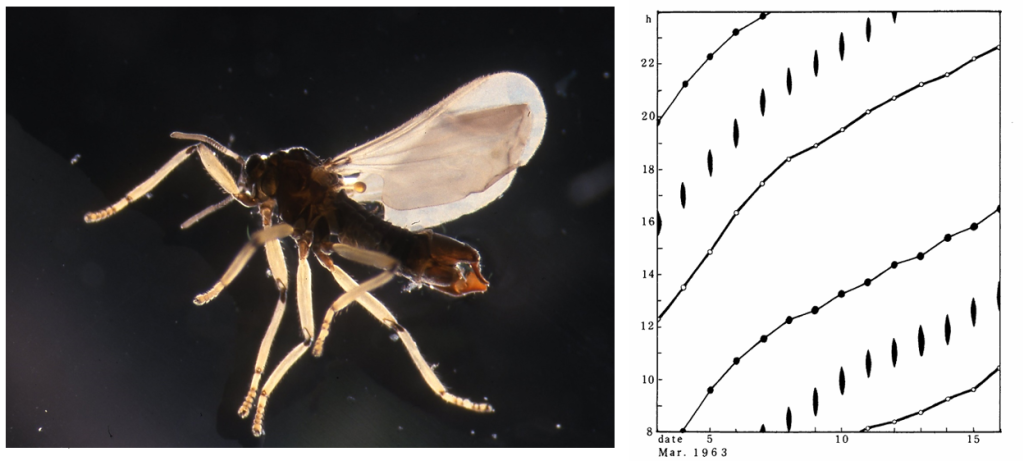
図11.(上)羽化したウミユスリカ(オス)。(右)ウミユスリカの一種。ともに西表島の海岸で採集。
種名(species)については,今は亡き橋本碩(ひろし)氏に聞いてみないと何とも言えない。橋本碩氏が奄美大島で観察したのはClunio takahashii(アマミウミユスリカ)だったので,西表島に分布しているのも同じ種かも知れない。時計細胞(潮汐時計と月周時計)は眼柄神経節のどこかにある。また,地質時代のいつ時計細胞が出現したか?羽化にともなって眼柄神経節や脳で発現する遺伝子を調べることで,時計細胞の位置が特定できるのではないだろうか?
私たちも瀬戸内海でウミユスリカ(Clunio tsushimensisと思う)の羽化を観察したことがある。ウミユスリカの羽化は,昼と夜の干潮の潮位(tide height)と深く関係している。昼間に潮が引く夏は、羽化は日中に起きる。夜間によく潮が引く冬は,羽化は夜間に起きる、春と秋には潮位が似たような高さになり,その時には羽化は昼と夜の一日2回起きる。橋本碩氏の野外観察は記載的な段階にあるが,現在(2026)でも羽化の観察は色あせていない。ウミユスリカの羽化のタイミングは,今後は遺伝子発現の面から研究できると思う。

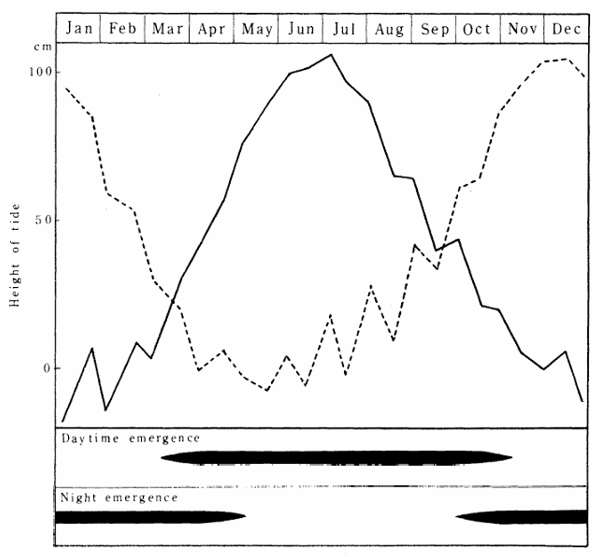
図12.アマミウミユスリカの羽化と潮位(tide height)との関係。
ウミユスリカの羽化リズム(semilunar rhythm)の研究では,ドイツのDietrich Neuman氏が有名である。しかし,橋本碩氏も負けず劣らずいい仕事をしている。残念ながら,日本人は一般にアメリカやヨーロッパの研究者に対して強いコンプレックスをお持ちの方が多い。要するに,自分自身で客観的な判断ができない人たちが多いということ。自分自身で判断すると権威者の大変な怒りを買うため,自分の判断は控えているのかも知れない。私は,権威者を上手くおだてるのが苦手である。すぐに怒らせてしまう。権威者は根が義理人情に厚い人たちなので,情に掉させばどこに流されるかわからない。そんな人たちの貴重なご教示はもう要らない。私は最後まで一人で戦いたい。何と言われようとも・・・。生真面目な性格は,こういう時(人から批判されたとき)に危機に陥る。図12(左と右)は,Hashimoto (1976)より転載した。
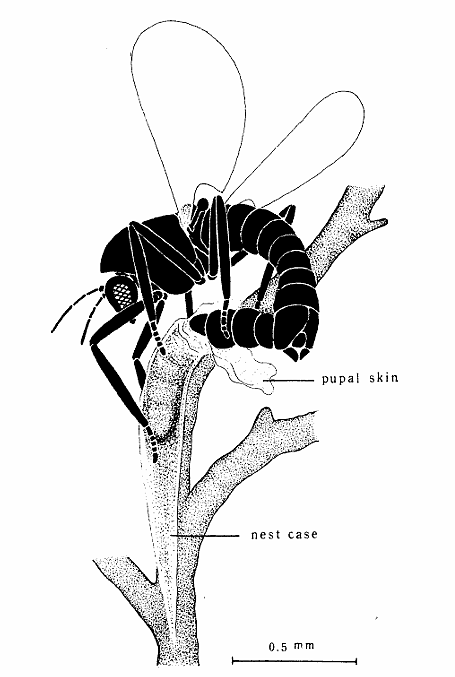
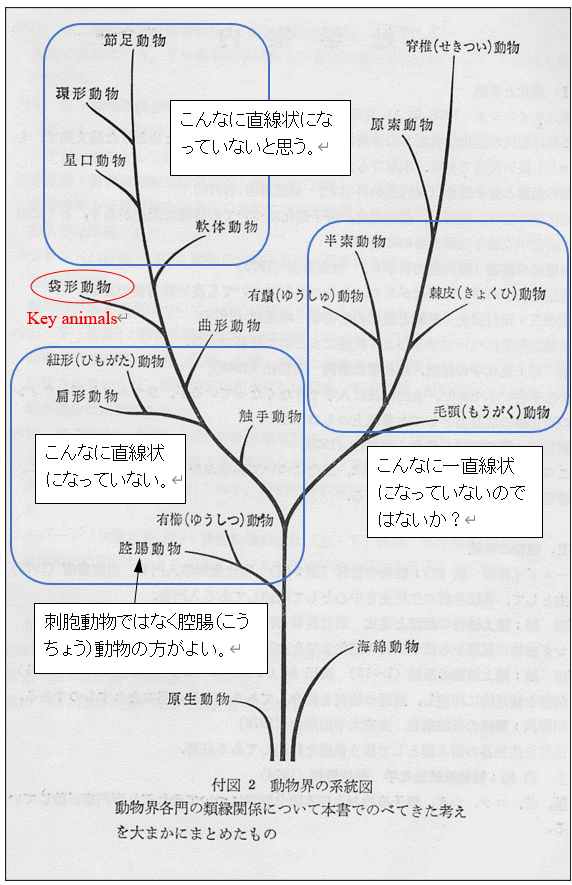
図13. 無脊椎動物の起源と進化(ひとつの仮説)
動物界(Animalia)は多くの門(phylum)で構成されている。それぞれのphylumの生物群(taxa)は,先カンブリア代(10億年前)に単細胞生物(真核細胞)から生じたと予想されている。このような単細胞生物(Phylum Protista)は,もっと原始的な原核生物(単細胞)が,共生して誕生したと考えられている。
大事なことは,原核細胞では単細胞が維持されたが,真核細胞では多細胞化が起きたということだ。共生(symbiosis)といっても,これ以上原核細胞を細胞膜の中に取り込めなくなったという事情があったのかもしれない。多細胞化は,いったん離れて生活する生物(真核細胞)が互いに身を寄せ合って生じたのではないだろう。生物の性質として,いったん離れてしまうと自己と非自己が認識されるようになり,非自己を認識する因子(まだ解明されていない)が取り除かれない限り,2つの真核細胞が接着することは不可能になる。自己と非自己の認識は,単細胞であれ多細胞であれ,個体性(individuality)の確立を意味する。
単細胞生物は,細胞内にエネルギーの取り込みが増加したときに,分裂してエネルギーを分配する。原生生物は分裂した後に互いに離れてしまうが,多細胞生物(後生動物ともいう)では,分裂した後に互いに身を寄せ合って,それぞれの細胞に機能分化が生じた可能性がある。接着によって,それぞれの細胞に機能分化が生じることの「必然性」はまだうまく説明できない。
図13にみられるように,確かに進化における系統(phylogeny)はあると思う。系統の生じる実験的証拠もいくらか持っている。一方では,進化における系統が高等な体制(organization)に向けて,こんなに直線上に配列されるのかという疑問が残る。脊椎動物の進化であれば,有袋類の適応放散にも,有胎盤類の適応放散にもそんなに強い方向性があるという証拠は全く出ていない。
無脊椎動物であれば,袋型動物門(Aschelminthes)はkey groupになるかも知れない。袋型動物の起源は不明である。袋型動物の共通の特徴は「偽体腔」であるが,これは体節構造(segmental structure)と関係しているように思う。体腔(偽体腔と真体腔)の出現は,偽体節構造から真体節構造への進化と関係しているかもしれない。偽体節や真体節を持った袋型動物がカンブリア紀に出現し,急速な勢いで多様化し,それらから多くの種類の無脊椎動物が進化したのだろう。軟体動物のネオピリナは体節構造を持つ。ネオピリナの体節構造は二次的に獲得されたとされているが,そうではないだろう。ネオピリナは,体節を持った袋型動物として最初に出現したのだろう。(図13は,山田・西田・丸山(1981)から転載)

図14.宮古島のサンゴ礁原。2024年7月3日。2024年は,6月17日から20日まで石垣島,6月21日から30日まで西表島,7月1日から4日まで宮古島,7月5日から12日まで沖縄本島に滞在して調査を行った。1971年は宮古島には寄らなかった。西表島以外は,長期滞在して研究する施設がない。
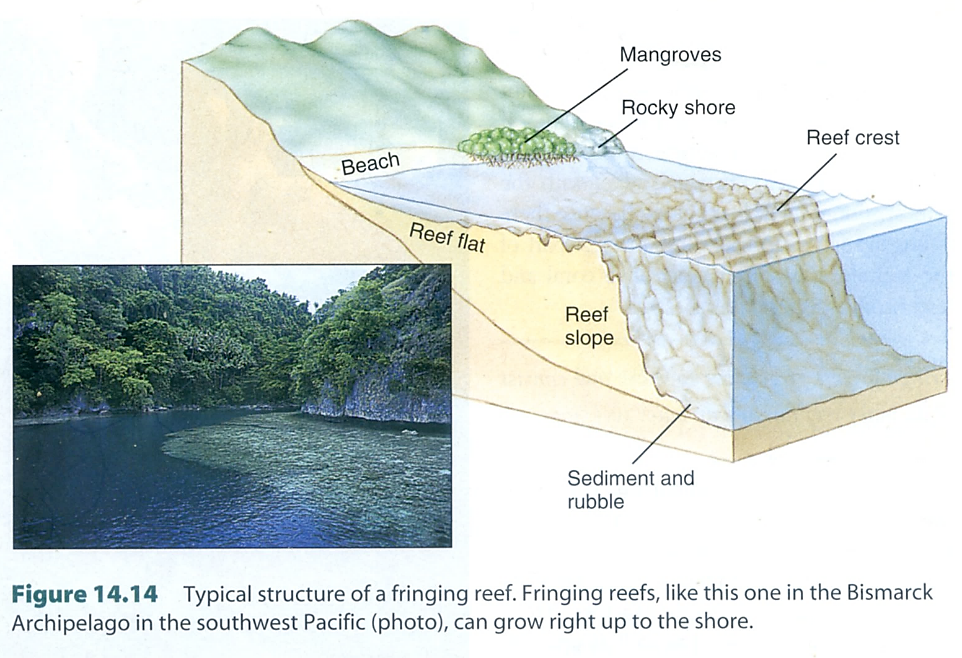
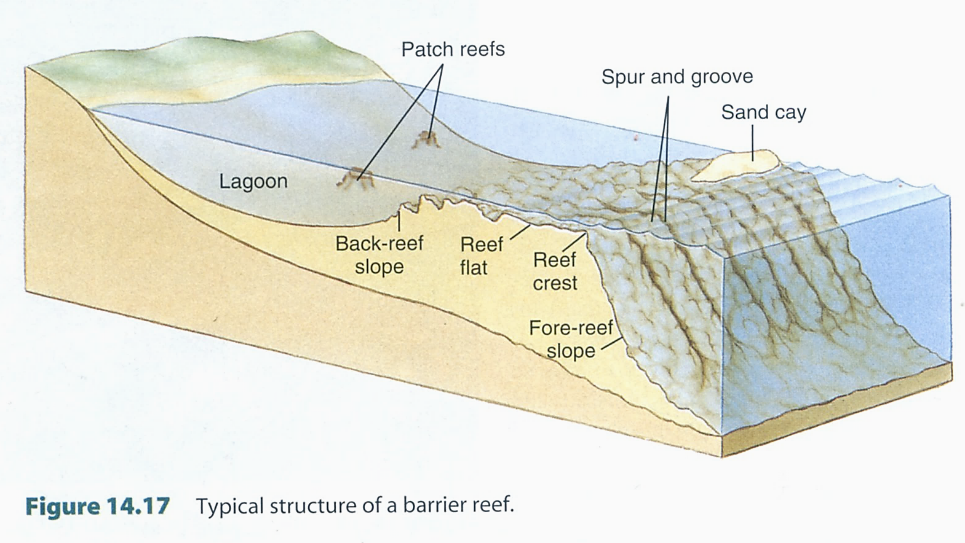
図15.熱帯域の海岸に見られるサンゴ礁原の構造。
世界のサンゴ礁は,3つのタイプに分類されている。陸上に近いのが裾礁(fringing reef),外洋に向けて発達した礁原を持つタイプが堡礁(barrier reef),そして南太平洋に見られる陸上が消失したタイプが環礁(atoll)になる。fringing reefは構造(structure)から定義されているが,barrier reef は機能(function)から定義されている。環礁(atoll)は構造からの定義。二重の定義になっているので,サンゴ礁のタイプがうまく分類できていない。
たとえば上の図(Figure 14.14)では,サンゴ礁原の構造と景観が一致していないので,fringing reefとbarrier reefの違いがよくわからない。サンゴの研究は進んでいるのに,琉球弧のサンゴ礁の構造が上の3つのタイプのどれに該当するかを述べている人はいないかもしれない。
裾礁にしても堡礁にしても,サンゴ礁の形成年代が異なっているだけの話であろう。形成年代が新しければ裾礁になるだろうし,古くなるにつれて堡礁になってゆくだろう。南太平洋では,島の誕生とともにサンゴ礁の形成も始まる。何万年もすれば島々は海中に没してしまうことが多いので,海面近くには発達しつつあるサンゴ礁が残って構造的には環礁となる。結局,琉球列島に見られるサンゴ礁のタイプは,裾礁といってもよいし,堡礁とも言える。
サンゴ礁の構造で重要な特徴がある。琉球列島のサンゴ礁の構造は上の図(Figure 14.14)に近い。特徴はreef flat(礁原)が浅いこと。大潮(spring tide)の干潮時(low tide)には,サンゴ礁原の縁(白波が立っているところ)まで歩いて行けることも多い。一方,もっと南に行けばサンゴ礁原はさらに発達して陸上とサンゴ礁の縁には礁湖(lagoon)ができる。礁湖の深さは5mから10mにおよぶ場合もあるだろう。そうなると,サンゴ礁の縁(reef flatからreef crest)まではボートで行くことになる。
もうひとつ,琉球列島のサンゴ礁の特徴について述べておきたい。それは,琉球列島のサンゴ礁原は潮の干満がゆっくりと起きるため,波はおだやか。グアム島(海洋島)だと波が荒く,干満も急に起きるので十分注意。Castro and Huber (2005) から転写。
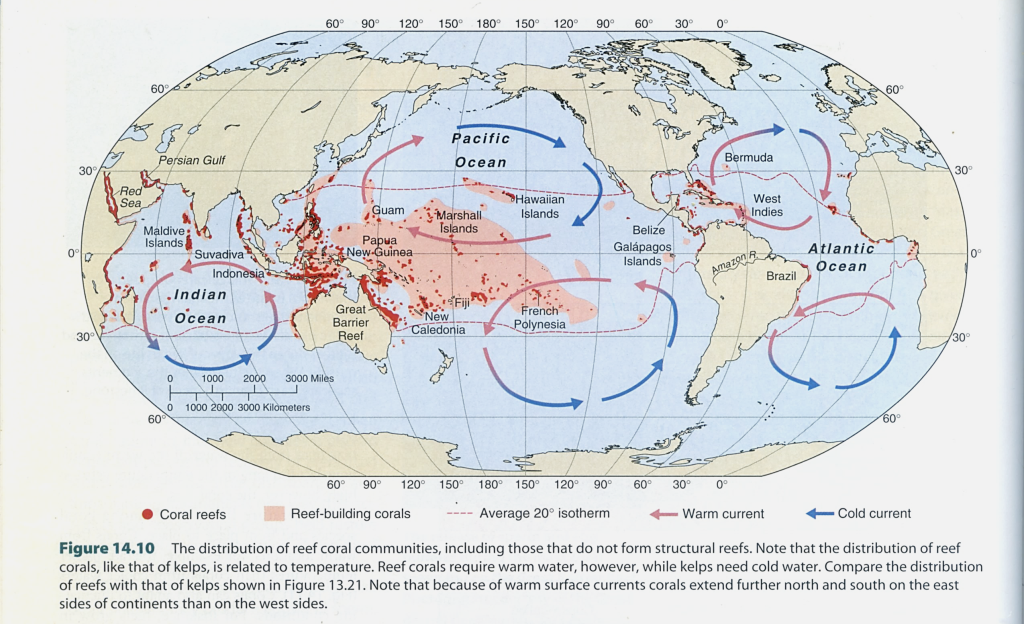
図16.世界のサンゴ礁の分布。サンゴ礁は,太平洋の熱帯域にある島々の周囲に発達するが,その中心はフィリッピン,ボルネオ,ニューギニアあたりにあるのかもしれない。礁を作るサンゴ(イシサンゴの仲間)の種分化がこれらの海域で始まったのかもしれない。そうだとすると,現在のサンゴ礁は新生代,それも一番新しい「第四紀」(260万年前から始まった)という可能性が浮上する。琉球列島の広大なサンゴ礁原は,島々の出現と同時に開始されただろう。でき始めてから1~2万年しか経っていないように思われる。一方,現在のサンゴ礁とはだいぶ感じの異なった礁(reef)は,中生代からあったという指摘がある。三畳紀末からジュラ紀にかけての十脚甲殻類の多様化(Triassic Marine Revolution)は,サンゴ礁の発達と関係するかもしれない。図はCastro and Huber (2005) から転写。

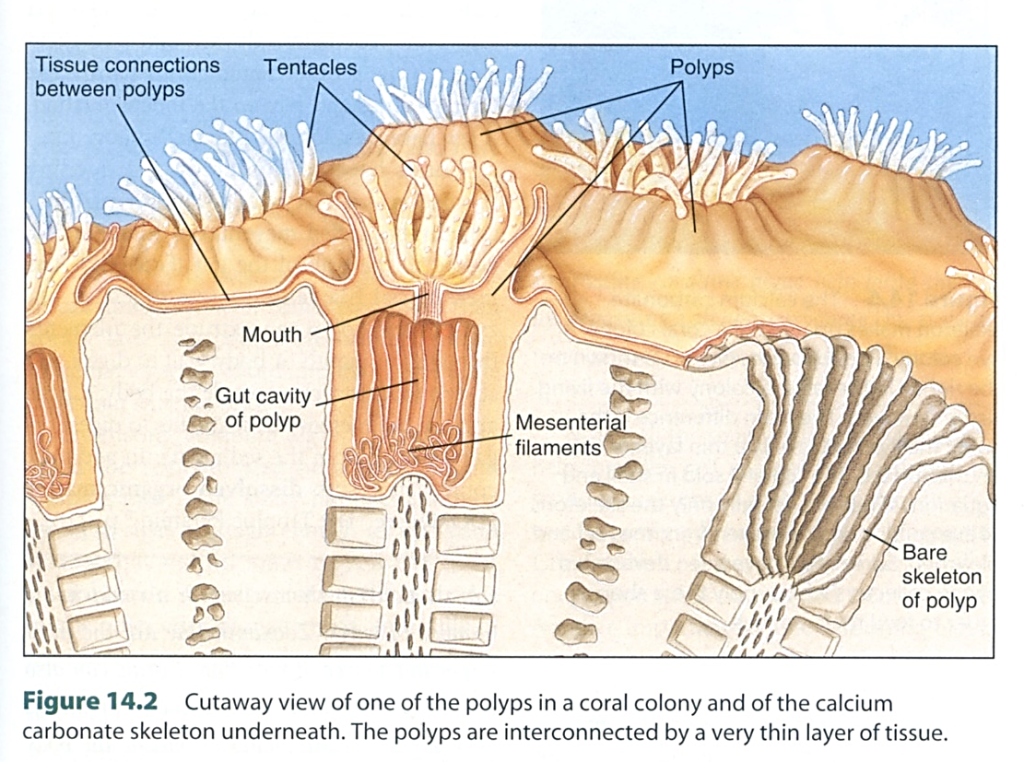
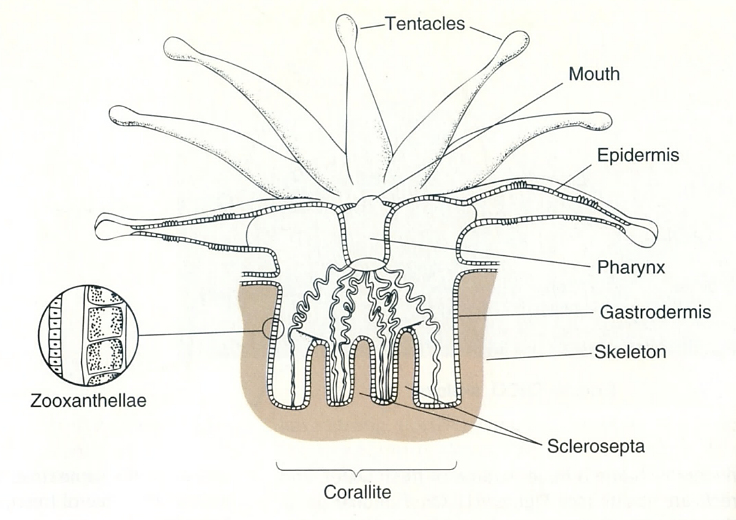
図17.(FIGURE 9.19) 熱帯域に分布する礁(reef)を作るサンゴ(主にイシサンゴ類)の種類数(属の数)。イシサンゴ類の種類数からみると,分布の中心はフィリッピン,ボルネオ,ニューギニアあたりにありそうだ。下の図(2つ)は,サンゴ塊を構成する個虫(individual worm)の形態を示す。サンゴ虫に取り込まれた動物プランクトンは,腸内で消化され,未消化物は再び口から排出されるのだろう。扁形動物の消化器系も似たような構造を持つ。割と原始的な構造を持つ多くの生物が,いわゆる高等な生物とともに地球上に生き残っているのは面白い。左図はCastro and Huber (2005)から, 右図はNybakken (2001)から転写。

図18.宮古島と伊良部島の間にある河口(estuary)に現れた泥干潟(mud tidal-flat)。宮古島と伊良部島は狭い入り江によって仕切られている。仕切りの部分は,満潮時(high tide)には入り江(inlet)となるが,干潮時(low tide)には干潟(tidal flat)ができる。潮が引くと泥干潟の上には,多くのカニ類が現れる。目の前には泥砂を引っ掻いた跡があるが,これはミナミコメツキガニが作った泥団子である。宮古島の泥干潟には,シオマネキ類が多いが,中でも繁殖期のルリマダラシオマネキはとんでもなくきれいな瑠璃色になる。撮影は2024年7月3日。写真左奥が海岸への出口。なお,宮古島周辺は琉球列島の他の海域に比べ海水の透明度が高い。

図19.水際に集結したヒメシオマネキの集団(宮古島Jul 3, 2024)。ヒメシオマネキは潮が満ちてくると泥の中に潜り,潮が引くと穴から出て泥干潟を徘徊する。徐々に水際に集まり,干潮時なると水際に沿ってヒメシオマネキの行列ができる。手前にいるのはミナミトビハゼ。シオマネキを全く恐れていない。

図20.多良間島のサンゴ礁原。陸上からサンゴ礁の縁まで150 mぐらいだろうか?現在(2026)多良間島には那覇空港から定期便(航空機)があると思う。

図21.石垣島東海岸のサンゴ礁原。石垣島の東側の海岸には広大なサンゴ礁原が形成されている。サンゴ礁原の種類は裾礁(fringing reef)なのか,堡礁(barrier reef)なのか,定義できていないと思う。ニューギニアとオーストラリアの間の海域に形成されるサンゴ礁をbarrier reefと定義するのであれば,日本の亜熱帯の島々に見られるサンゴ礁はfringing reefと呼ぶことになるが,日本の亜熱帯の島々のサンゴ礁も十分に「波消ブロック」としての機能を果たしていると思うが・・・。

図22.石垣空港に向けて降下している飛行機(JAL)。あと10秒もすれば着陸になる。奥の小山はガーラ岳。以前の石垣空港は,新空港より5 kmほど南西にあった。滑走路は短く,弾丸着陸が行われていた。着陸寸前にそっとシートベルトを外して置くと,着陸時の急ブレーキで体が起き上がって起立した状態になった。当然ながら,客室乗務員からものすごく怒られた。若い時はよくこんなことをして,あちこちで怒られていた。こういうところは紳士的な関口晃一氏とは違っていた。

図23.1971年の石垣港桟橋(11月に撮影)。
離島に荷物(泡盛の一升瓶)を運ぶために積み込み作業をしている。ひめゆり丸はこの桟橋に着いたと思う。なお,現在の離島桟橋(高速船)は100mほど離れた場所に移転したが,貨物船(「かりゆし・平成丸」とか「カーフェリー・ぱいかじ」は,古い方の桟橋から出ていると思う。東海丸や第三幸八丸も古い桟橋から出港したと思う。どこから乗船したかの記憶はない。
写真の手前にマリヤ牛乳の箱が置いてある。マリヤ牛乳もよく覚えている。株式会社マリヤ乳業は,今も営業しているみたいだ。
大学では,こんな記事(No. 3)を書くと必ず密告する者が現れる。大学を通して検閲を受けたら,記事はたちまち発禁になる。多くの大企業なども同様な事情があるのだろう。NPO法人として大学や研究所とは独立した組織を作ると自由に物が書け,気持ちが楽になる。
一方的に文句を言われっぱなしというのも困るが,そういう連中に文句を言い返すと泥沼にはまる。密告する者は,自分に勝てなかった人たちと思えばよい。ろくに戦わずして始めから負け犬になっている人たちは,どうにも救いようがない。傷をなめ合うこともできない。
それと,私は学会には入りたくない。例えば,日本動物学会は科学研究費の配分をめぐって権力者のまわりに集まる烏合の衆という感じを受ける。研究費の採択や職務の地位などでゴマをすると足元を見られ,権力者の手下にされる。最初は仕方がないが,いつまでもそんな関係を続けていると頭がおかしくなる。対等な関係で共同研究ができると良い。
<参考文献>
- Castro, P., and M.E. Huber (2005) Marine Biology. Fifth Edition. McGraw-Hill Higher Education. Boston.
- Ferl, R.J., and R.A Wallace (1996) Biology: The Realm of Life. Third Edition. HarperCollice Publishers, New York.
- 池田嘉平・稲葉明彦(1971) 日本動物解剖図説。森北出版。
- 木村政昭(1996) 琉球弧の第四紀古地理。地学雑誌105: 259-285.
- 木村資生(1988) 生物進化を考える。岩波新書。
- Hashimoto, H. (1964) Notes on Thalassomyia japonica from Ryukyu (Diptera, Chironomidae). Kontyu 32: 311-322.
- Hashimoto, H. (1965) Discovery of Clunio takahashii Tokunaga from Japan. Jap. J. Zool. 14 (3): 13-29.
- Hashimoto, H. (1968) Seasonal variation in the marine chironomid Clunio tsushimensis Tokunaga (Diptera). Ann. Zool. Fenn. 5: 41-48.
- Hashimoto, H. (1976)Non-biting midges of marine habitats (Diptera: Chironomidae). Pp. 377‒414. In: L. Cheng (ed.), Marine Insects. North-Holland Publishing Company, Amsterdam.
- 平野誉(2016) 化学発光と生物発光の基礎化学。 化学と教育 64(8): 376–379.
- 岩切晴二(1974, 1975) 数学精義 Ⅰ, Ⅱ, Ⅲ. 培風館。
- Martin, J.W., J. Olsen, and J.T. Høeg (2014) Atlas of Crustacean Larvae. Johns Hopkins University Press, Baltimore.
- Neuman,D. (1966) Die lunare und die tägliche Schlüpfperiodik der Mücke Clunio. Steuerung und Abstimmung auf die Gezeitenperiodik. Z. Vergl. Physiol. 53: 1-61.
- Nybakken, J.W. (2001) Marine Biology: An Ecological Approach. Fifth Edition. Benjamin Cummings. San Francisco.
- Oka, H. (1930) Morphologie und Ökologie von Clunio paciñcus Edwards. Zool. Jahrb. Abt. Syst. Ökol. Geogr. Tiere 59: 253-280.
- Oka, H., und H. Hashimoto (1959)Lunare Periodizität in der Fortpflanzung einer pazifischen Art von Clunio (Diptera, Chironomidae). Biol. Zentralbl. 78: 545-559.
- 近江谷克裕(2016) 発光プローブの開発を目指した発光生物の探索(2)。昆虫と自然51 (11): 34-36.
- Saigusa, M. (1981) Adaptive significance of a semilunar rhythm in the terrestrical crab Sesarma. Biol. Bull. 160: 311–321.
- Saigusa, M. (1982) Larval release rhythm coinciding with solar day abd tidal cycles in the terrestrial crab Sesarma – harmony with the semilunar timing and its adaptive significance. Biol. Bull. 162: 371–386
- Saigusa, M. (2001) Daily rhythms of emergence of small invertebrates inhabiting shallow subtidal zones: A comparative investigation at four locations in Japan. Eco. Res. 16: 1–28.
- Saigusa, M., and T. Akiyama (1995) The tidal rhythm of emergence, and the seasonal variation of this synchrony, in an intertidal midge. Biol. Bull. 188: 166–178.
- 関口晃一(1984) カブトガニの生物学。サイエンスハウス。
- 関口晃一(1991) カブトガニの不思議: 生きている化石は警告する 。岩波新書。
- 関口晃一(1994) カ ブ トガニ の適応戦略と種分化。化石 56: 7−50.
- 関口晃一(2008) いつの間にやら米寿を越えて。アサヒ印刷。
- 山崎柄根(2012) 関口晃一博士(1919–2012)を想う。日本動物分類学会誌(タクサ) 33: 1–3.
- コルバート E.H. (田隅本生 訳)1978. 脊椎動物の進化(上巻と下巻)。築地書館。
- 山村庄亮・長谷川宏司(2008)天然物化学‒海洋生物編‒。アイピーシー(理工学系の学術書出版社)。
- 山田真弓・西田誠・丸山工作(1981)進化系統学。裳華房。
執筆: 三枝誠行(NPO法人,生物多様性研究・教育プロジェクト,プロジェクト長。理学博士。)