
2025年4月25日(金)
ヨコヤアナジャコ(Upogebia yokoyai)とはどんな生物なのか?
河口域の川岸に行くと,干潮時(time of low tide)には泥干潟の表面に空いている小さな穴をたくさん見ることができる。小穴を作った張本人はアナジャコやスナモグリであることが多い。アナジャコの特徴としては,穴の出口にマウンド(塚)を作らないことである。一方,出口に大小さまざまなマウンドを作る種類は,スナモグリ(全部ではない),ハサミシャコエビ,オキナワアナジャコ(こちらは巨大なマウンドを作る)である。スナモグリはすべての種がマウンドを作ることはなさそうだ。
アナジャコ類(Upogebiidae)は,スナモグリ,ハサミシャコエビ,オキナワナジャコも含めて起源は古く,中生代の三畳紀と思われる。テチス海の広大な内湾の海底には,アカザエビやロブスターの祖先であるAstacidea(ザリガニ下目)の十脚甲殻類(decapod crustaceans)が繁栄していた。これらの一部は海岸に上がってきて,生活するようになったのだろう。中生代には陸上では植物(シダ類や裸子植物)繁栄し,大小さまざまな河川から河口や海岸に有機物が溜まり,それを餌とする十脚甲殻類が進化したと思われる。ザリガニ(Astacidaeでよいと思う)は,陸上の淡水域にまで進出したが,アナジャコやスナモグリの祖先は,海岸や河口域の砂泥底を住みかとしていたのではなかろうか?スナモグリは三畳紀(2億5千万年前から2億130万年前)から化石が出ているが,アナジャコについては新生代,それも第四紀(258万8千年前から現在まで)あたりと思われる。アナジャコが2億年の間どこでどうしていたのかはわからないが,おそらくAstacidea(ザリガニ下目)の化石の中に含められてしまっているのだろう。原著論文は執筆中の段階ではあるが,アナジャコ・スナモグリ・オキナワアナジャコ・ハサミシャコエビの祖先がアメリカザリガニ(Procambarus clarkii)の祖先と近い系統関係にあることを示す明確な証拠(担卵毛の微細構造)がある。
今までは,動物の形態を見て俺はアナジャコの分類の専門家だから,これは何々という種と判断できる。・・・みたいなことで研究が進んできたが,それでは子供のケンカが続くだけのことである。まずはdescriptiveなデータ(客観的なデータ)をよく見て,そこからものを考え,議論する方向に進めて行きたい。動物のスケッチでは主観的な判断が優先され,ケンカは収まらない。客観的なデータとしてはミトコンドリアDNAや核DNAの分析があるだろう。写真(photograph)も十分に客観的なデータになりうる。

図1.イシアナジャコの生息場所(沖縄本島・屋我地の海岸)。破線の中にある泥岩の中に穴を掘っている(図2左の写真)。この海岸に干潮時に露出する泥岩は硬く,逃げ足も速いので採集はかなり難しい。イシアナジャコは,糸満市の名城ビーチの泥岩に住むUpogebia rupicola(図2の中央と右)ではないかと思われる。和名については,オキナワイシアナジャコとでもしたらどうだろうか?生物多様性科学の分野では,分類学と共通である必要はない。2024年7月上旬に撮影。

図 2.Upogebia rupicola。左の写真は屋我地島産の♂。中央と右がそれぞれ名城ビーチ産の♂と♀。Upogebia のグループは雌雄の形態的な違いが大きい。アナジャコ類は,メスの方は腹部第一体節の付属肢(appendage)が生えているが,オスの方は付属肢がない。しかしながら,オスにもかかわらず腹部第一体節の付属肢が見られる個体がいる。さらにオスにもかかわらず,胸部第 3 歩脚の付け根に「メス」の生殖口がみられる個体がいる。ただしオスに見られる「メスの生殖口」は機能はしていない。笠岡湾の干潟に住む大型の種類(Upogebia major)では,オスなのにもかかわらず「卵巣」が見られる。もちろんオスなので精巣も見られ,こちらは機能している。オスの卵巣はメスに比べて小さく,卵成熟は起きない。(輸卵管も生殖口につながっていない。)十脚甲殻類では性決定の仕組みはよくわかっていないが,環境ホルモンによってオスがメス化する可能性はないだろう。いずれにせよ,オスとメスでは形態に大きな違いがあり,別種としないよう注意されたい。
形態による種の認定
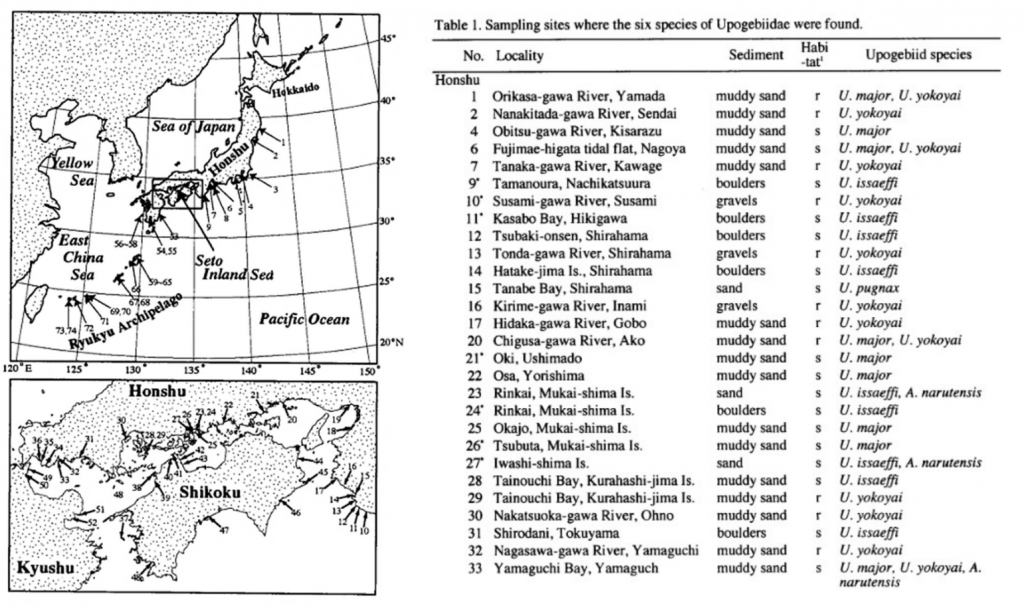

図 3.本州,四国,九州,および琉球弧に分布するアナジャコ。古典分類学に基づく種名の決定。表は Itani (2004)から転載。
Itani (2004)によれば,琉球列島に分布しているアナジャコ(ヒメアナジャコ)は 3 種類(Upogebia yokoyai, U. carinicauda, U. pugnax)が生息している。しかしながら,ヒメアナジャコ類は形態的によく似ているために,U. pugnax を除き形態だけでは種を判別することが難しい。私たちも琉球弧においてアナジャコを採集し,まず形態で分類を試みたが,Itani (2004)の分類とは一致しなかった。分子系統解析を加えて考察すると,琉球列島には次の 5 種類が分布していることが判明した。
Genotype B,Genotype C,Genotype D,Upogebia rupicola(図 2),および Upogebia pugnax の 5 種類が区別できる。なお Upogebia pugnaxは宮古島の河口で採集。生きているときの形態と体色を見れば,分子系統解析をしなくても,他種との区別は容易である。
本州・四国・九州には別な種類のヒメアナジャコが 3 種類(GenotypeA, Genotype E, Genotype F)分布する。なお,Itani (2004)のいうUpogebia yokoyai は Genotype A か E か F のどれに該当するかは不明
である。模式標本を取り寄せられたところで,形態学的な観察からでは,分類学者との溝は埋まらない。Upogebia yokoyai という種は,古典分類学では存在しても,生物多様性科学では認知できない種になっている。分子系統学的解析に加えて,形態学的にも明瞭な違いがあるので各 genotype(A~F)はそれぞれ別種とみなしてよい。
<琉球列島に分布>
Upogebia ishigaki (genotype D); U. Kariyushi(genotype B); U. okinawa-jima (genotype C). U. rupicola と U. pugnax も。
<本州・四国・九州に分布>
Upogebia nihonnkai (genotype A); U. Kodono (genotype E?); U. hatsukaichi (genotype F?)
なお,本州・四国・九州に生息する大型のアナジャコ(U. major) 等については今のところ変更がない。
分子系統学的解析によるgenotypeの検出と形態の比較による種の判定
私たちも中国地方の瀬戸内海側と日本海側・四国・九州の沿岸域の泥干潟をまわって,ヨコヤアナジャコ(Upogebia yokoyai)を採集した。採集は,奄美大島を始めとして,沖縄本島・宮古島・石垣島・西表島でも行われた。琉球列島の島々をめぐるヨコヤアナジャコの採集行脚はすごく楽しい。
実験動物こそが生物学の研究に最も貢献すると思っている人たちは,野外で行う採集や実験は観光旅行と同じように考えている。そういう人たちは,自然科学の研究はこうあるべきだとか,人間とはこういう風に生きるものだとかという倫理観念(先入観)が異常に強い。自分たちの脳の中に埋め込まれた先入観に少しでも外れるような行動をすると,恐ろしい剣幕で怒り出す。文系・理系ともに研究者にはよく見られる人格のタイプである。
こういうの(自称人格者)に従っていると,心理的なストレスが急上昇して,精神的に弱い人だと「うつ病」にかかる。逆に「信者」にされてしまい,他人に対して同じ行動をとるようになるので,注意が必要である。日頃「自分の意見が言える」という環境を整えておく必要があるだろう。うつ病や狂信者にされてから上司のハラスメントが認定されたところで,病気になった自分を元に戻すことは難しい。
・・・という話の詳細は別の機会に譲るとしたい。この記事では,広範囲にわたる採集標本について,姜奉廷さんが濱口氏の研究室で遺伝子解析(mtDNAのシークエンス)を行った。結果としてgenotype Aからgenotype Fまでの6つの遺伝子型の個体群があることが判明した(論文は執筆中)。これらのうちgenotype Aとgenotype Eに関しては,生活史のパターンの分析から互いに別種である可能性が強く示唆された(Yamasaki et al. 2010)。
私たちが2008年に行った採集の結果,genotype Aは西日本の中国地方,四国,九州の河口域に広く分布することが分かった。またgenotype Eは日本列島の河口域に局地的に分布することも判明した。両者が混在する干潟もあった。一方,genotype B, genotype C, およびgenotype Dの遺伝子型は琉球列島に分布していることが分かった。それぞれのgenotypeは,同じ干潟に生息していることが多かった。もしそれぞれのgenotypeがひとつの種であった場合には,同じ干潟に生息しているのであれば,当然交雑が起こり,ひとつの干潟には単一のgenotypeしか残らないはずである。
遺伝子解析で種に対応する違いがあるのだったら,それぞれの個体群には形態の違いも検出できるかもしれない。琉球列島では,実際に形態が異なるヨコヤアナジャコのグループがいるか調べてみようという目的で石垣島,西表島,宮古島,沖縄本島を回った。宮古島のマングローブでもヨコヤアナジャコ(・・・と今までは思っていたが,Upogebia pugnaxという別種であることが判明)が採集できるので,この島も調査の対象にした。
奄美大島にはgenotype Bとgenotype Cが分布することがわかっており,genotype Bは石垣島で,genotype Cは沖縄本島でも記録されている。それぞれの遺伝子型の形態を比較し,種(species)と認定できる違いがあるかを検討した。
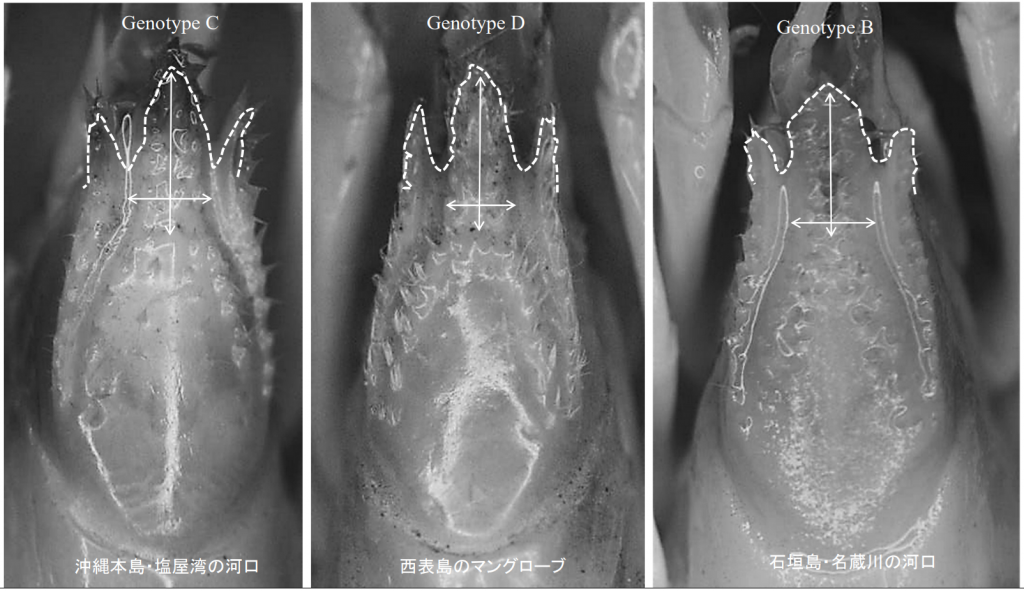
図4.琉球弧に分布するヒメアナジャコ(Upogebia属)の3種類のgenotype。Genotype Cは沖縄本島でいちばん多くみられるタイプ。塩屋湾と本記事で紹介する丁間川のヒメアナジャコはすべてgenotype Cであった。Genotype Dは西表島のマングローブに広く分布する。 Genotype Bは,石垣島の名蔵川の河口で採集されているが,名蔵川の干潟は2~3種類のヒメアナジャコが混じっていた。琉球弧のヒメアナジャコは,本州や四国に分布する種類(図5)と違って,頭部の両端にある「襟(えり)」が長いのが特徴である。
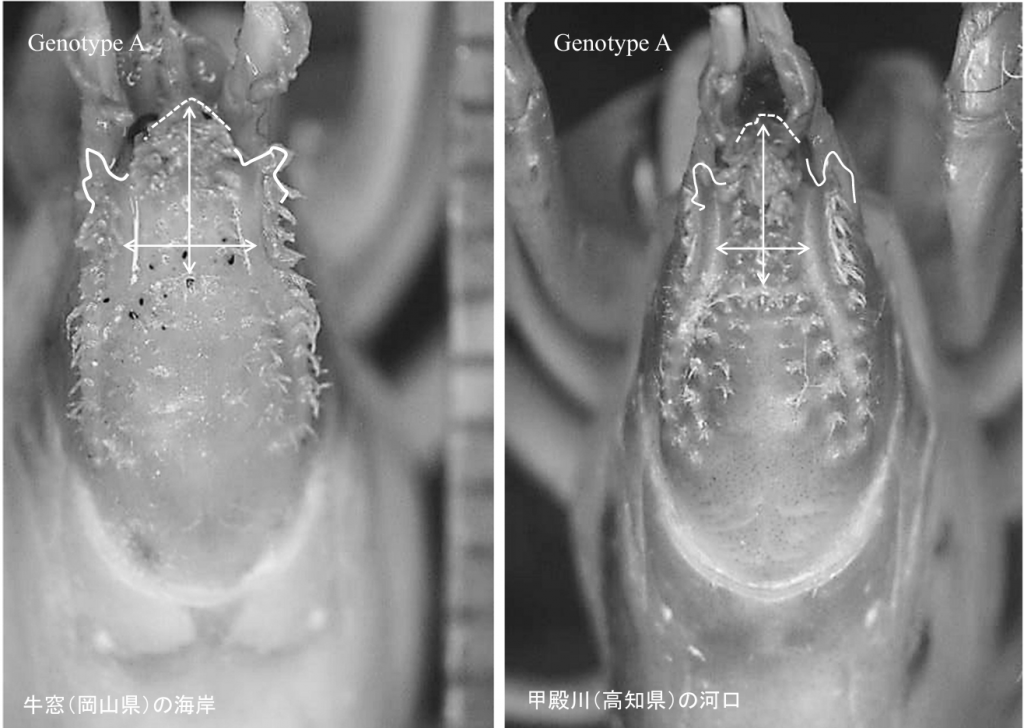
図5.本州や四国に分布するヒメアナジャコ。左の写真は,岡山県牛窓(うしまど)の海岸で採集した個体,右は高知県春野町・甲殿川の河口で採集した個体。 ともにgenotype A に属する個体である。しかし,genotype Aは,Itani (2004)の言うUpogebia yokoyaiかはわからない。Itani (2004)には種の判別に必要な写真は掲載しておらず,種類に関しては伊谷氏本人しか知りえない情報をもとに判定している。専門家に相談しなければ,研究が進まないという時代ではなくなっていると思う。
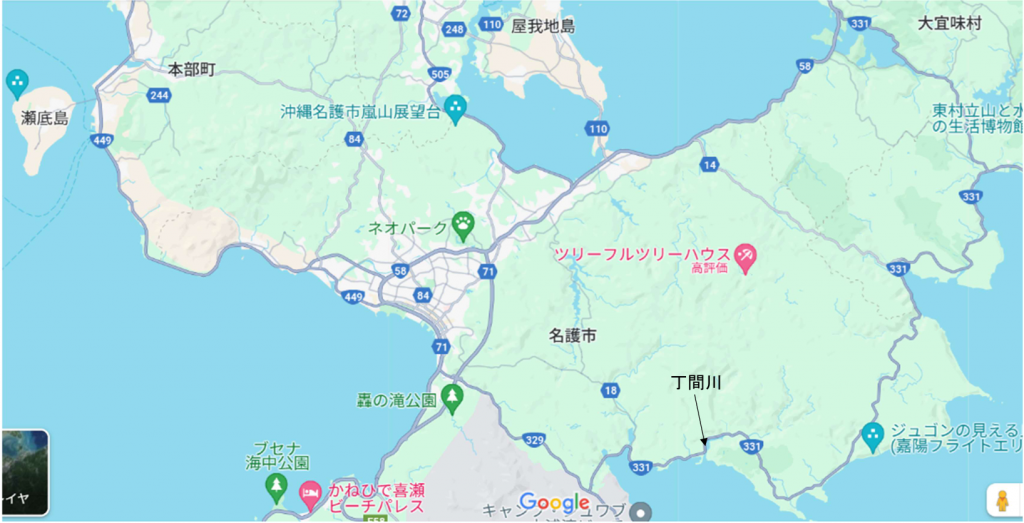
図6.丁間川(名護市)の場所。沖縄本島の中部から南部にかけては,川がほとんどなく,干潟のできる河口は少ない。私がいつも行くのは,名護市から奥(沖縄本島最北端の集落)までの範囲に限られる。沖縄本島における採集は,いつも瀬底島にある琉球大学熱帯生物圏研究センターの瀬底研究施設(以前の理学部付属瀬底臨海実験所)に滞在して行われる。瀬底島は,以前は本部港から船で往復したが,現在は瀬底大橋があるので本部半島から車で往復できる。
丁間川に行くコースはいくつかある。以下に私が使った道を紹介する。まず研究施設から瀬底大橋を渡り,本部町の方に行く。本部町から県道84号を通り,伊豆味まで行き,そこから左折して県道123号に入り,海岸沿いの505号に出る。海岸沿いを走ると羽地に出る。羽地から国道58号で源河まで行き,右折して県道14 号に入る。県道14号を行くと東海岸にある県道331号に出る。県道331号を南に下れば20分ほどで丁間川に出る。帰りは早く行きたければ,331号から県道329 号を使い名護に出て,それから449号を通って研究施設に戻るコースがある。帰りはBIG本部店(スーパー)でビールと夕食を買った。
丁間川での採集は,2024年7月9日に行われた。以下,丁間川と周辺の風景,および丁間川で採集されたヒメアナジャコ(genotype C)の写真を掲載した。
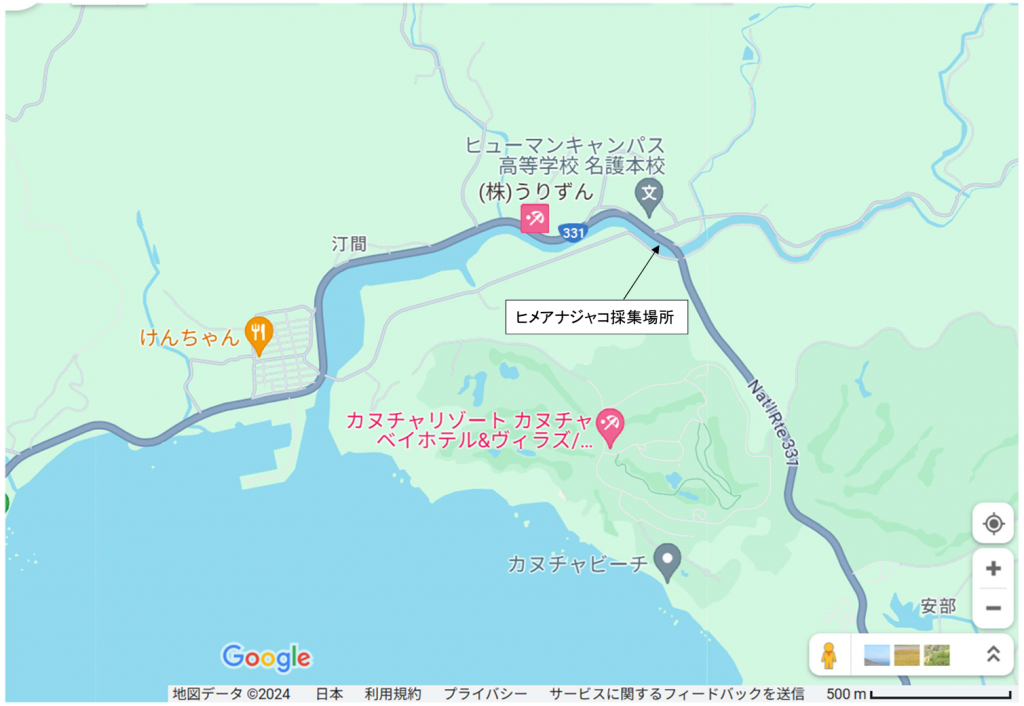
図7.丁間川におけるヒメアナジャコ(Upogebia sp.)の採集場所。Itani (2004)らの言うヨコヤアナジャコ(Upogebia yokoyai)は,分子系統解析を加えて分類したgenotype のどれに対応するかわからない。形態的特徴のみでは,アナジャコ類の種を判定するのは不可能である。分類学者は採集されたアナジャコを見て,これは何々という種だと断定するだろうが,自分の経験に基づいた主観的な判断で行われる。これでは現代生物学の進歩から取り残されるだけである。

図8.丁間川は,20年以上前に訪れたことがある。その時と2024年7月9日に見た風景は全く同じものだった。20数年前もこの場所でレンタカーを止めて石段を下り,全く同じところ(干潟)でアナジャコを採集した。20年前には何という種かもわからなかったが,20数年を経た現在では,ここに住むアナジャコはヒメアナジャコのGenotype Cに属する種類(和名はオキナワヒメアナジャコ)で,沖縄本島の北部一帯の河口に広く分布していることがわかっている。

図 9.オキナワヒメアナジャコの巣穴。ヒメアナジャコの巣穴(入り口)は,海岸で潮が引いた時(干潮時)に川底をのぞくと見ることができる。巣穴の入り口は 2 つあり,この場所だと5~6 cmほどの深さでひとつになる。丁間川のこの辺り(図7)では軟泥層は浅く,巣穴はせいぜい30 cmほどの深さだろう。

図10.丁間川の干潟(tidal flat)。この辺り(図7)の川底の底質は小石が多く,ヒメアナジャコは深い穴を作ることができないだろう。干潟の川底を30 cmも掘れば,ヒメアナジャコはたくさん捕れる。ヒメアナジャコは,ホンアナジャコ(Upogebia major)と形態はよく似ているが,生息場所が分かれている。ホンアナジャコは,大きな干潟の軟泥層に深い巣穴を掘っているが,ヒメアナジャコは干潟の縁の小石混じりの川底に巣穴を掘って住んでいる。季節にもよるが,ホンアナジャコを採集するのは重労働である。ヒメアナジャコは,マングローブに堆積する軟泥層では採集に苦労するが,この場所(図7)では容易に採集できた。沖縄本島の河川は,海岸の砂が海岸への出口に堆積し「河口閉塞」が起きることが多い(特に西側の海岸)。そのため河口域の塩分濃度(salinity)は淡水のそれに近くなり,アナジャコは住めない。沖縄本島ではアナジャコを採集できる干潟は少ない。本部半島周辺の河口には生息場所が多い。

図11.丁間川の河口に住むヒメアナジャコ。遺伝子型で言うとgenotype Cに該当する。の学名は,Upogebia okinawa-jimaとしてみた。分類学では,種名には「i」「ensis」「nicus」などつけることが多い。しかし,どんな種類かを知るためには,種の特徴や採集場所を種名の中に入れる方がよい。また,ヒメアナジャコのそれぞれのgenotypeは,いわゆる亜種(subspecies)の違いではなく,れっきとした種の違いである。多様性プロジェクトの出版する原著論文でも,分類学の論文と同じように,そこで採集されたのはどんな種かを記述することは重要な作業である。分類学では権威者の鶴の一声で種名が決定されるが,生物多様性科学では客観的な証拠(例えば写真を使っての形態比較とか,分子系統学の解析結果)の上に立って種が決定される。つまり自称専門家の方々のご意見をお伺いしなくても,分類学や系統進化学の豊富な知識と技術があれば,誰でも学問に参入できる。専門家にお伺いを立てないと学問ができないという悪習はもう終わりにしたい。

図12.オキナワヒメアナジャコ(Upogebia okinawa-jima)。ヒメアナジャコの仲間は,頭部背面にある「シールド」と呼ばれる上皮の硬い部分の形態を比較することで種を判定できる(図4と図5)。そのほかの特徴,例えば鋏脚の「長節」(merus)の表面に生える棘の有無や数なども種を分ける参考にはあるが,棘の数は鋏脚の自切(autotomy)が起きてから再生(regenerate)すると変わることがある。この写真(図12)は,RICOH WG50を使って撮影されているが,この程度のresolutionではヒメアナジャコであることすらも十分にわからない。顕微鏡下での頭部撮影の写真でないと種を見分けることは難しい。Itani (2004)が,このような標本の種名をどのよう判定したかは不明であるが,私たちの研究とはあまりに接点が少ないために議論をしようがない。

図13.オキナワヒメアナジャコ(斜め横から撮影)。ヒメアナジャコはアルコール固定すると頭胸部が反り上がる。このような状態で真上からスケッチすると,全然違った形態になる。スケッチはまだ写真の技術がない時代に導入された描画技術であり,写真技術の発達した現代では出番が減少している,と私は思う。

図14.真上から見た時のオキナワヒメアナジャコ(♀)。EOS 7Dを使って撮影した。EOS 7Dだと撮影台があればもっとよい写真が撮れるだろうが,2024年はあまりに荷物が多すぎたので,顕微鏡・顕微鏡撮影装置・撮影架台は西表島から岡山に送り返してしまった。沖縄本島にはgenotype Cが広く分布するが,この写真では,genotype Cであると判断できない。この個体を分類学者に見てもらったら,皆U. yokoyaiと判断するだろう。平野優理子さんも,ヨコヤアナジャコは1種類という先入観があっただろうから,この写真を見せたらUpogebia yokoyaiと判断すると思う。
しかし,分子系統学的知見とシールドの形態比較という2つのスタンダードで見ると,丁間川に生息するヒメアナジャコはgenotype Cとするのが妥当だという結論になった。それぞれのgenotypeは,いくつかの証拠から「種レベルの違い」とみなすことができるという結論から,日本列島に分布するヒメアナジャコはUpogebia rupicola と U. pugnaxを含めて計 8種類ほどに分類できた。一方では,昔ながらの視点でヒメアナジャコを分類する方法も残っている訳だから,互いに議論すればよいだろうという指摘もあると思う。しかし,研究の視点がもともと違っているので,妥協点を見出すことは難しい。そんな努力をするのであれば,古典分類学とは方針の異なる「生物多様性科学」という新しい研究分野を設立し,その中で生物の分類・系統・進化を研究するのが良いだろうと思った。だから,私の系統進化学には,過去の分類学者の仕事を紹介し,その功績をたたえるという内容は全く含まれていない。批判する気はないが,周囲はそう思うのだろう。

図 15.丁間川の河口。写真左側には海岸がある。右側は上流。橋(丁間橋?)は海岸から 50m も離れていないところに架かっている。橋の奥(上流)の泥干潟にはアナジャコ類の巣穴がたくさん見られるが,橋の手前(海岸)には全く見られない。住んでいてもよさそうな気がするが・・・。

図16.丁間川の河口(ほぼ海岸)。干潮時に川底に露出する泥岩は硬く,イシアナジャコの巣穴も見当たらない。転石の下には別な種の巣穴があるかもしれない。

図17.橋から100mほど上流にある土手の前にできた泥干潟(mud tidal flat)。この辺り(海岸から150ほど上流)になると,泥干潟にはたくさんの穴が見られる。大きい穴はヒメシオマネキ,小さな穴はヒメアナジャコが作ったのだろう。その他,ハサミシャコエビもいると思う。この場所は穴が深く,ヒメアナジャコは採集できなかった。

図 18.干潟の生物を観察している親子。観察しているのは,ハクセンシオマネキと思われる。ハクセンシオマネキは,潮間帯の砂干潟に多く見られる。干潮時になっても干潟に出るときには,小さい子供の場合には必ず親がついて行く必要がある。泥干潟には危険な場所が多く,ちょっと目を離したすきに深みにはまっていたり,カキ殻に触ってケガをすることも多い。しかし,小さいころからこんな場所で遊び慣れていると,どこに危険があるか見ただけでわかるようになる。

図19.丁間川の近くにある集落(場所不明)。春の夕方には,ふ化直前の卵(胚)をたくさん抱えたベンケイガニ(Sesarma intermedium)が山から下ってきそうだ。さすがに7月上旬では繁殖シーズンは終わっているので,次の年の繁殖シーズンまでベンケイガニは山の中にいるはずだ。

図20.丁間川の近くの集落で見かけた飲料水の自動販売機。沖縄本島には,たしかコカ・コーラのマークが入った自販機とLifeguardのロゴが入った自販機の2種類が置かれていると思う。Lifeguardの方はCoca colaに比べて安いが,どれも味は今ひとつである。私はいつも「日本サイダー」というドリンク(100円)を買っていた。余談ではあるが,私の書いた記事は分類学者や生態学者にとってはおよそ面白くないだろう。いわゆる専門家を自称する人たちは,見ることはないと思う。私の対象とするところは,野生生物の多様性に多少とも関心のある一般の方々である。ドリンクにも多様性がある,・・・なんてことも知っていただければと思う。
<参考文献>
- Futuyma, D.J. (1998) Evolutionary Biology. Third Edition. Sinauer Associates, Inc. Massachusetts.
- Castro, P., and M.E. Huber (2005) Marine Biology. Fifth Edition. McGraw Hill Higher Education. Boston.
- Itani, G. (2004) Distribution of intertidal upogebiid shrimp (Crustacea: Decapoda: Thalassinidea) in Japan. Contr. Biol. Lab. Kyoto Univ. 29: 383-399.
- Hirano, Y., and M. Saigusa (2008) Description of the male of Upogebia miyakei from Ryukyu Islands, Japan (Decapoda: Thalassinidea: Upogebiidae). J. Mar. Biol. Assoc. UK 88: 125‒131. DOI: https://doi.org/10.1017/S0025315408000234
- Karasawa, H., C.E. Schweitzer, and R.M. Feldmann (2013) Phylogeny and systematics of extant and extinct lobsters. J. Crust, Biol. 33: 78–123. https://doi.org/10.1163/1937240X-00002111.
- Nanri, T., M. Fukushige, J.P. Ubaldo, B.-J. Kang, N. Masunari, Y. Takada, M. Hatakeyama and M. Saigusa (2010) Occurrence of abnormal sexual dimorphic structures in the gonochoristic crustacean, Upogebia major (Thalassinidea: Decapoda), inhabiting mud tidal flats in Japan. J. Mar. Biol. Assoc. UK 91: 1049‒1057. DOI: https://doi.org/10.1017/S0025315410001712
- Ngoc-Hon N. (1994) Notes on some Indo-Pacific Upogebiidae with descriptions of four new species (Crustacea: Thalassinidea). Mem. Queensland Mus. 35: 193‒216.
- Nybakken, J.W. (2001) Marine Biology. An Ecological Approach. Fifth Edition. Benjamin Cummings, San Francisco.
- Oka, T.B. (1941) Oocyte-like cells in the ovarial part of the testis of Gebia major. Journal of the Faculty of Science, Imperial University of Tokyo, Section IV 5, 265–289.
- Yamasaki, M., T. Nanri, S. Taguchi, Y. Takada, and M. Saigusa (2010) Latitudinal and local variations of the life history characteristics of the
- thalassinidean decapod, Upogebia yokoyai: a hypothesis based on trophic conditions. Est. Coast. Shelf Sci. 87: 346‒356.
- 山田真弓・西田誠・丸山工作(1981)進化系統学。裳華房。
<撮影と執筆の基本情報>
撮影と執筆:三枝誠行(生物多様性研究・教育プロジェクト常任理事)
撮影機材:野外ではCanon EOS 7Dと RICOH WG-50。種類の判別は顕微鏡撮影装置(Zeiss Stemi 2000CにCマウントアダプターを介してCanon EOS 7Dをつけた)を利用。