
1.Introduction
私たちの身近にいて,頻繁に食卓に上がるエビ類やカニ類は,分類系統学上で言えば節足動物門(Phylum Arthropoda)の甲殻綱(Class Crustacea)に属する。・・・と言っても,甲殻綱自体もたくさんの亜綱(subclass)から成り,カシラエビ亜綱(Cephalocarida),鰓脚亜綱(Branchiopoda;ホウエンエビ,カブトエビ,ミジンコ),橈脚(とうきゃく)亜綱;カラヌス),蔓脚(まんきゃく)亜綱(Cirripedia;カメノテやフジツボ),それに軟甲亜綱(Malacostraca)が含まれる。
さらに,軟甲亜綱にも多くの目(order)が含まれる。コノハエビ目(Nebaliacea),アミ目(Mysidacea),クマ目(Cumacea),タナイス目(Tanaidacea),等脚目(Isopoda),端脚目(Amphipoda),オキアミ目(Euphausiacea),それにエビ・カニ類の十脚目(Decapoda)が含まれる。このように,エビ・カニ類は,十脚目の中のほんの1グループに過ぎない。まあ,何と数多くの種類の甲殻類がいるものだと感心される方もおられると思うが,すべてを知らなければ学問ができないという訳ではない。お目当ての生物とその周辺に注目していればよい。
地球上では,30数億年前に最初の生命が誕生した。以来,絶滅した種類も半端なく多いが,全体としては種類数は増加した。だから今日の地球の姿があるのだろう。しかし,この先は今のように急激な速度で種類数が増加することはないだろう。そして,全体として種類数が減少してゆく時代(終生代)がやってくるに違いない。どうあがいても,来るものは来る「時」が来る。
さてと,節足動物門の甲殻目のエビ・カニ類では,体の構造(structure)に大きな特徴がある。節足動物は,体の構造が多くの体節(segment)からできている。そして,それぞれの体節に対応して,体節の側面から1対の付属肢(pleopod)が生えている。各体節の外皮は,石灰化したプレートで覆われていて,体節ごとにプレートの境界溝(boundary)があることだ。
甲殻類の中で一番原始的とされるカシラエビを見ると,頭部(1体節),胸部(顎脚と歩脚)の8体節,および腹部の11体節が区別され,各体節のプレートは明確な溝(くびれ)で仕分けされている。
ところが,十脚甲殻類の根鰓(こんさい)亜目(Dendrobranchiata)に属するクルマエビでは,胸部の各体節(第1顎脚から第3顎脚と,第1歩脚から第5歩脚)のプレートが複雑に融合していて,上から見るといくつ体節があるかわからないほど変形している(図1)。腹部の体節はそのまま残り,全部で7体節あることがわかる。
抱卵亜目の十脚甲殻類で面白いところは,胸部の第3顎脚のプレートと第1歩脚のプレートの間に明確な境界線(boundary)が現れることである。この境界線(浅い溝)を「頸溝」(cervical groove)と呼ぶ人もいるが(椎野 1964),顎脚・歩脚プレート境界溝,またはMPプレート境界線と呼ぶ方が,より強い印象を与えるだろう。
繰り返しになるが,甲殻類では頭部から尾部に至るまで,それぞれの体節の外皮(exoskeleton)となるプレートに対して,1対の付属肢(pleopod)が生えている。最初のうちは,すべての付属肢が歩行(locomotion)に用いられていただろう。しかし,生物が進化するに連れて,付属肢に機能分化(functional differentiation)が生じたと考えられる。腹部の7体節では,付属肢は遊泳のために特化した構造(遊泳肢)に変化する一方,頭部や胸部の方では第1から第3付属肢が顎脚(maxilliped)に変化した。
付属肢の機能分化にともない,特に胸部では各体節のプレートに大きな変化が現れた。つまり,顎脚は大あご(mandible)の後方に強く圧縮され,エサの捕獲と捕食のために使われるようになった。ところが,胸部内壁には大きな胃があるために,外皮(プレート)の方は,元の形態を維持して胸部の容積を確保する必要に迫られたのだろう。
結局,何が起きたかというと,胸部の3つの顎脚プレートは複雑な形に融合したのと,そのあおりを受けて顎脚に続く5対の歩脚プレートも複雑に融合した構造になったと考えられる。しかしながら,第3顎脚のプレートと第1歩脚(鋏脚)のプレートの間には明瞭な溝(groove)が残ってしまったのだろう。
私は,何年にもわたってこの溝(境界線)の由来を考えてきた。そして,MPプレート境界線は,顎脚と歩脚に対応するプレートの仕切り線(溝)でないかという結論に至った。幸い,MPプレートは,石灰化された甲羅(carapace)の上に生じるので,化石として残る。私の学問の大きなメリットは,現生の十脚類の形態学と古生物学との接点を見出したことである。
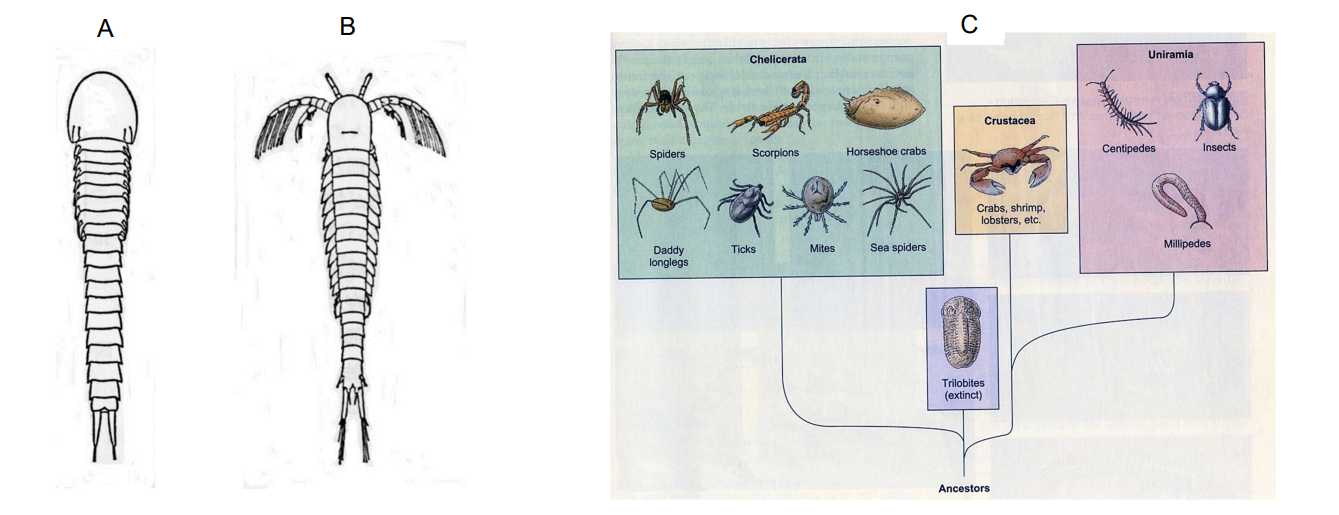
図 1.甲殻類の起源と進化。A:Hutchinsoniella macracantha(背面)。ハチンソニエラは現生の種。B:Lepidocaris(デボン紀の化石)。C:甲殻類の起源。Trilobita は三葉虫。三葉虫は海底を歩行していただろうから,基本的に各体節の付属肢は歩脚だったと思われる。図 1C の Crustaceaの 6 本脚は,どう見てもおかしい。(A と B は,山田・西田・丸山「進化系統学」から転載。C は,Ferl・Wallace「Biology」から転載。
2.撮影と執筆の基本情報
<サンプルの採集地> それぞれの写真の説明文の中に,採集場所と年月日を記した。
<採集者と記事の執筆者> 三枝誠行(NPO 法人,生物多様性研究・教育プロジェクト常任理事)
<Key words> 十脚甲殻類,根鰓亜目,抱卵亜目,アカザエビ,アナエビ類(Axiidea),ロブスター(Homarus),アカザエビ,西表島(舟浮)
3.参考文献
・山田真弓・西田誠・丸山工作(1981)進化系統学。裳華房。
・Ferl, R.J. and R.A, Wallace. 1996. Biology: The Realm of Life. Harper Collins College Publishers.
・Fraaije, R.H.B. (2014) Diverse Late Jurassic anomuran assemblages from the Swabian Alb and evolutionary history of paguroids based on carapace morphology.
N. Jb. Geol. Paläont. Abh. 273/2: 121-145. Stuttgart.
・Fraaije, R.H.B., Ban Vakel, B.W.M., Jagt, J.W.M., Charbonnier, S., Schweigert, Garcia, G., and Valentine, X. (2022) The evolution of hermit crabs (Crustacea,
Decapoda, Paguroidea) on the basis of carapace morphology: a state-of-the-art-report. Geodiversitas 2022 44 (1): 1-16.
・内田亨(1965)動物系統分類の基礎。北隆館。
・椎野季雄(1964)節足動物(I)総説・甲殻類。動物系統分類学 7(上)。中山書店。
・Williams, A.B. 1995. Taxonomy and evolution. In: Factor, J.R. (ed.) Biology of the Lobster Homarus americanus. Academic Press.
・池田嘉平・稲葉明彦(監修)1971.日本動物解剖図説。森北出版。
4.研究方法と引用文献
私の学問は,甲殻類の歩脚の発現に関係する遺伝子の種類とか発現様式に関する最新の研究を追いかける方向とは,真逆ではないにしても,同じ方向にはない。著名なジャーナルの編集者は,流行りの研究を推奨しているけれども,私は,流行りものについて行く中で,自分を認められたいとは思っていない。正直に言えば,権威者の持ついい加減さや信念をありがたく頂戴すると,結局は自分の身を滅ぼす結果になる。
私には,十脚甲殻類の種分化・系統・進化について,自分なりに育んできた興味・関心がある。残る人生の中で,十脚甲殻類の種分化・系統・進化について,可能な実験や実証的な証拠をもとにして論じてみたい。流行りの研究でないとインパクトファクターの高いジャーナルに掲載される可能性は低くなるが,エビデンス(evidence)は不可欠なので,原著論文の出版は続けなければならない。
具体的には,中生代三畳紀のサンゴ礁の地形や構造と,中生代における十脚目(order)・下目(infraorder)・種(species)の多様化のプロセスを知りたい。権威者なる人々に尋ねても,いい加減な返事しか返ってこないか,初めから拒否されるかのどちらかである。ならば,自分の育んできた基礎学力と,精いっぱい努力するという意欲を前面に出して,新しい道を歩んでゆくのがよいだろう。最新の情報はインターネットで得ることができる。インターネットの記述(例えば Wikipedia)は,岩波生物学辞典や理化学辞典に比べて正確さは劣るが,そこは自分の基礎学力で補えばよい。何事も権威に頼らなければ先に進めない時代は,終焉を迎えているのではないだろうか。
岩波生物学辞典や理化学辞典は,図鑑や英和大辞典(研究社)と同様に,図書館や学校に飾りとして置かれるだけで,利用はされないだろう。
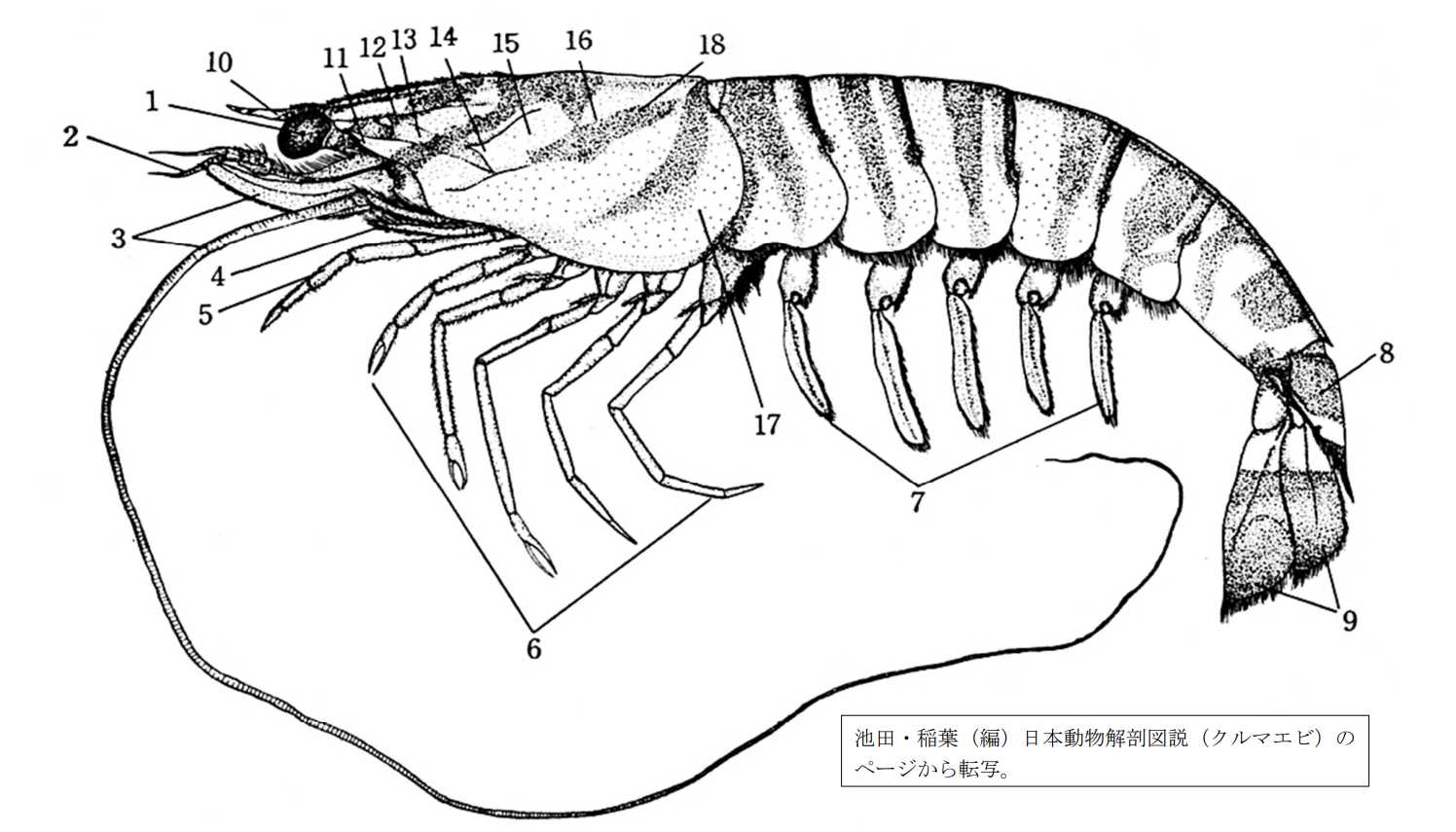
図 2.クルマエビ(根鰓亜目)の体の構造。頭部,胸部(第 1 顎脚~第 3 顎脚),および腹部(第 1 歩脚~第 5 歩脚)から成る。つまり,頭部は 1 体節,胸部は 8 体節,腹部は 7 体節から成り,それぞれの体節の外皮はプレート構造になっている。十脚甲殻類が進化するに当たり,頭部と腹部は問題ないが,胸部については 3 対の顎脚(maxilliped)が大あごの周囲に圧縮されてしまったことである。一方,胸部のプレートは元の形態を維持しようとしたのだが,腹部のように各体節を維持するという訳には行かず,各プレート間での融合が生じたのだろう。融合の名残を正確にトレースすることは難しいが,十脚類では腹部と胸部の境(boundary)に加えて,第 3 顎脚と第 1 歩脚(鋏脚)の仕切り(groove)が残っているのが大きな特徴である。

図 3.クルマエビの一種(種名は不明)の頭部と胸部。2007 年,5 月に大潮の干潮時に西表島(干立)で撮影。クルマエビの受精卵は,メス親の腹部にある担卵毛(ovigerous setae)に付着せず,受精直後に海中にばら撒かれる。根鰓亜目は,十脚甲殻類の中で一番原始的と考えられる。地質時代のいつごろ出現したかは不明であるが,古生代と言っている人が多い。私は,中生代の三畳紀にテチス海の内湾で,アミ目の一部が底生化して生じたのではないか,と考えている。頭部は,左右の眼柄の基部を結んだあたりまで。それより後は顎脚のプレート,さらに歩脚のプレートが続く。顎脚のプレート境界がどのあたりにあるのか不明だが,私の予想では白い楕円の中あたりにあると思われる。MP プレート境界溝に関しては,根鰓亜目では不明瞭である。

図 4.クルマエビ類の頭部にある凶悪なトゲ(rostrum)。クルマエビ類は,潮が引いているときには,サンゴ礁の細かいサンゴ礫の中に身を隠している。潮が上げてくると(rising tide)サンゴ礫から身を乗り出し,満潮時(high tide)になると,海面をぴょんぴょん勢い良く飛び跳ねる。捕食者が捕まえようとすると,凶悪なトゲの一撃がある。サンゴ礫の中に身をひそめるか,サンゴ塊の隙間に潜む十脚類の頭部には鋭いとげのある種類が多い。

図 5.アカザエビ(Metanephrops japonicus)。土佐湾の底引網で獲れる。クルマエビと同様に,頭部先端にはトゲ(rostrum)が生えているが,アカザエビでは「根ほり」のような形状をしている。「根ほり」の後端は,MP プレート境界線の付近で閉じている。各プレートは強く癒合しているため,外皮全体が非常にフラットな形状をしている。アカザエビやロブスターのもう一つ大きな特徴は,胸部が扁平ではなく,円柱状(cylindrical)の構造になっていることである。アカザエビの化石は三畳紀の終わりごろから出ているだろう。なお,胸部右側の膨らみは,エビヤドリムシの寄生を示す。アカザエビは,高級食材として出荷されるが,スーパーの魚屋さん(高知でも)で見かけることはない。味は,ロブスターやザリガニとよく似ているのではなかろうか?十脚甲殻類の筋肉とか中腸腺(ミソ)は,プリン体の権化みたいなところなので,食べる気はしない。高知ではサガミアカザエビと並んでまだ網にかかっているが,相模湾では乱獲によってほぼ水揚げゼロという状態になっていると思う。日本列島周辺の深海(150m~300m)には,アカザエビ,サガミアカザエビ,ミナミアカザエビが分布する。ヨーロッパに分布するのは,Nephrops norvegicus。形状はやはりアカザエビ。

図 6.シロオビアカアナエビ(Axiopusis serratofrons)(抱卵個体)。2007 年 5 月 17 日舟浮(西表島)奥の浜で採集。私は長いことアナエビ類(サンゴ礁ロブスター)に興味を持っていたが,系統と進化については解決の糸口を見つけることができなかった。最近胸部の体節のプレート(外皮)の構造から化石を分類する研究(ドイツのグループ)を知り,アナエビ類にも適用した。すると,アナエビ類とアカザエビには,共通する特徴があることが分かった。アナエビにも明瞭な MP プレート境界溝があり,アカザエビのように「コ」の字型から,「V」字型あるいは「U」字型に変化しているのがわかる。

図 7.シロオビアカアナエビの生息する浜。2007 年 5 月 17 日。この浜は,西表島の白浜から定期船に乗り舟浮(ふなうき)まで行き,舟浮の集落から徒歩で裏側のイダの浜に出る。イダの浜からは,海岸沿い(大岩がごろごろしている)を 30 分ほど西に歩くと着く。このあたりの岩は柔らかい砂岩(泥岩か?)で,潮間帯は少しばかり泥浜の状況を呈している。外洋に面するこの浜では,大潮の干潮時になると泥岩や砂岩が露出するが,1 時間もすれば潮が上げて海面下に沈む。アナエビ類を採集するには時間がかかる。アナエビ類はこんな潮間帯にいるかもしれないが,採集するのはかなり難しい。
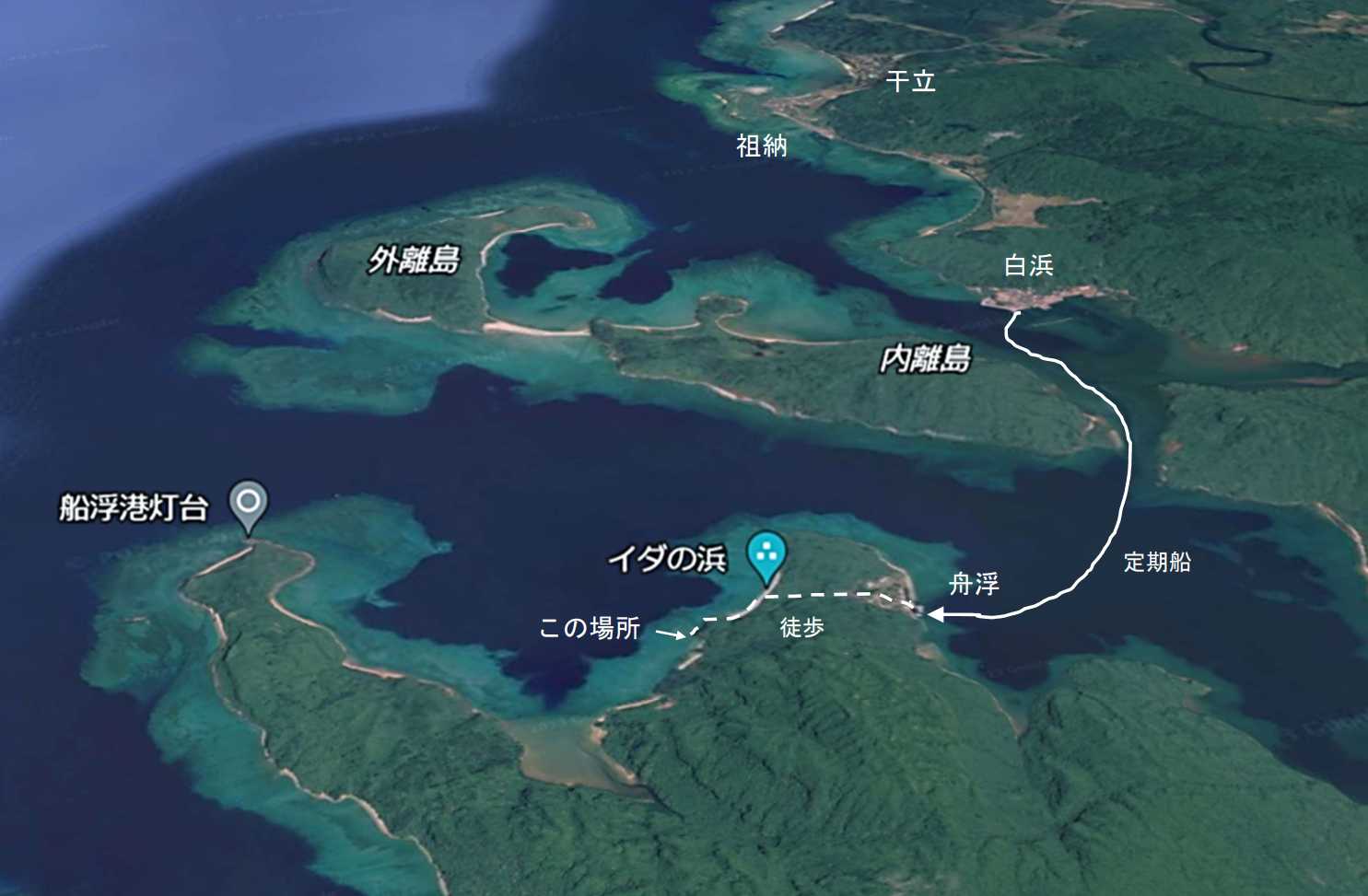
図 8.シロオビアカアナエビの採集場所。白浜から定期船で舟浮まで行き,そこから徒歩でイダの浜に出てから海岸沿いを歩く。岩の上を上ったり下りたりを何回か繰り返すので落ちないように注意。採集場所は崖になっているので,石や岩が落ちてくる危険性がある。崖の真下で仕事をしないよう。

図 9.シロオビアカアナエビの生息場所。シロオビアカアナエビは,サンゴ礫の中に巣穴を作ってペアで住んでいる。砂岩の縁を注意深く掘り進めると(深さは 10 cm もなかったように思う),30 分から 1 時間ぐらいで採集できることがある。なお,砂岩は硬く,侵入している種類はない。

図 10.イダの浜の奥にあるシロオビアカアナエビの生息地(大潮の干潮時)。平たい泥岩の上に打ち上げられた石灰岩と崖から落ちてきた砂岩が散乱している。石を取り除いたり,石の隙間を掘ると十脚甲殻類を採集することができるが,すぐ潮が上げてくるので,ゆっくり採集している時間はない。

図 11.イダの浜のさらに奥にある浜。海岸の露頭は砂岩。ここから先(網取方面)は,泥が流れ込むような浜はないので,行っていない。舟浮,あるいはかつて舟浮に住んでいた子供たちの中で,ここまで来た者はどのぐらいいるのだろうか?後ろは崖。地震があって津波が来れば,逃げ場がない。

図 12.干立(西表島)の西側海岸での採集。泥岩の周囲に堆積したサンゴ礫を掘ると,胸部が白く,腹部は赤色のスナモグリ(エラゲスナモグリ)がよく出てくる。アナエビ類はなかなか捕れない。2007 年 5 月,大潮の干潮時。干立の西側の海岸も崖になっている。崖の真下に荷物を置かないように。

図 13.アナエビ類の採集。 2007 年 5 月。西表島干立(ほしだて)西側の潮間帯。大潮の干潮時。とにかくサンゴ塊の周囲のサンゴ礫をがさがさとやっては,金網ですくうことを繰り返す。だいぶばててしまって,お尻は海水につかっている。1 日数匹しか採集できないことも多い。

図 14.シロオビアオアナエビ(Axiopusis pacificus)。 2007 年 5 月。西表島干立(ほしだて)西側の潮間帯。属(genus)はシロオビアカアナエビと同じだろう。学名(種名)は既に記載されているかもしれないが,分類学者に相談すると,標本は返してくれないは,種名は勝手に記載されるはで,とんでもないことになる。分類学者には,私の方から何かお願いすることはない。私は学会の規約(内規)などに頼るよりも,法律に頼って研究をしたい。

図 15.アカマダラアナエビ(Axiopusis iriomotensis)。 2007 年 5 月,西表島干立(ほしだて)西側の潮間帯で採集。属(genus)はシロオビアカアナエビと同じだろう。学名(種名)は既に記載されているかもしれない。アカマダラアナエビは,今までに 1 匹しか採集されていない十思う。アナエビ類の標本についても,当面分類学者の方々にご利用いただくことは難しい。私の研究が終わったならば,しかるべき手続きを踏んで利用可能になるだろう。

図 16.御畳瀬(高知)の底引網で採集された魚介類。深さは,200m~300m と思う。食卓に上がる種類は非常に少ないが,深海にすむ海産生物に興味を持っているごく少数の人々にとっては,狂喜するほど懐かしい写真だろう。きっとお宝があるはずだ。それも 1 つや 2 つだけでなく,初めて見るお宝がたくさん入っているかもしれない。お宝を期待して仕分け作業に入るのだが,とにかく時間がかかる。トロ箱一つにつき,仕分けは平均 1 時間かかる。

図 17.御畳瀬の底引網で採集された十脚甲殻類。底引網の深さは,200m~300m と思う。トロ箱から十脚甲殻類のみより分けた。この時(2019 年 12 月9 日)には,コシオリエビが大量に入っていた。多くはオオコシオリエビ(Cervimunida princeps)だが,チュウコシオリエビ類(Munida)も何種類か入っていた。ところで,コシオリエビ類は,何でオオコシオリエビだけ違う属名(Cervimunida)なのか?Munida ではまずいのだろうか?
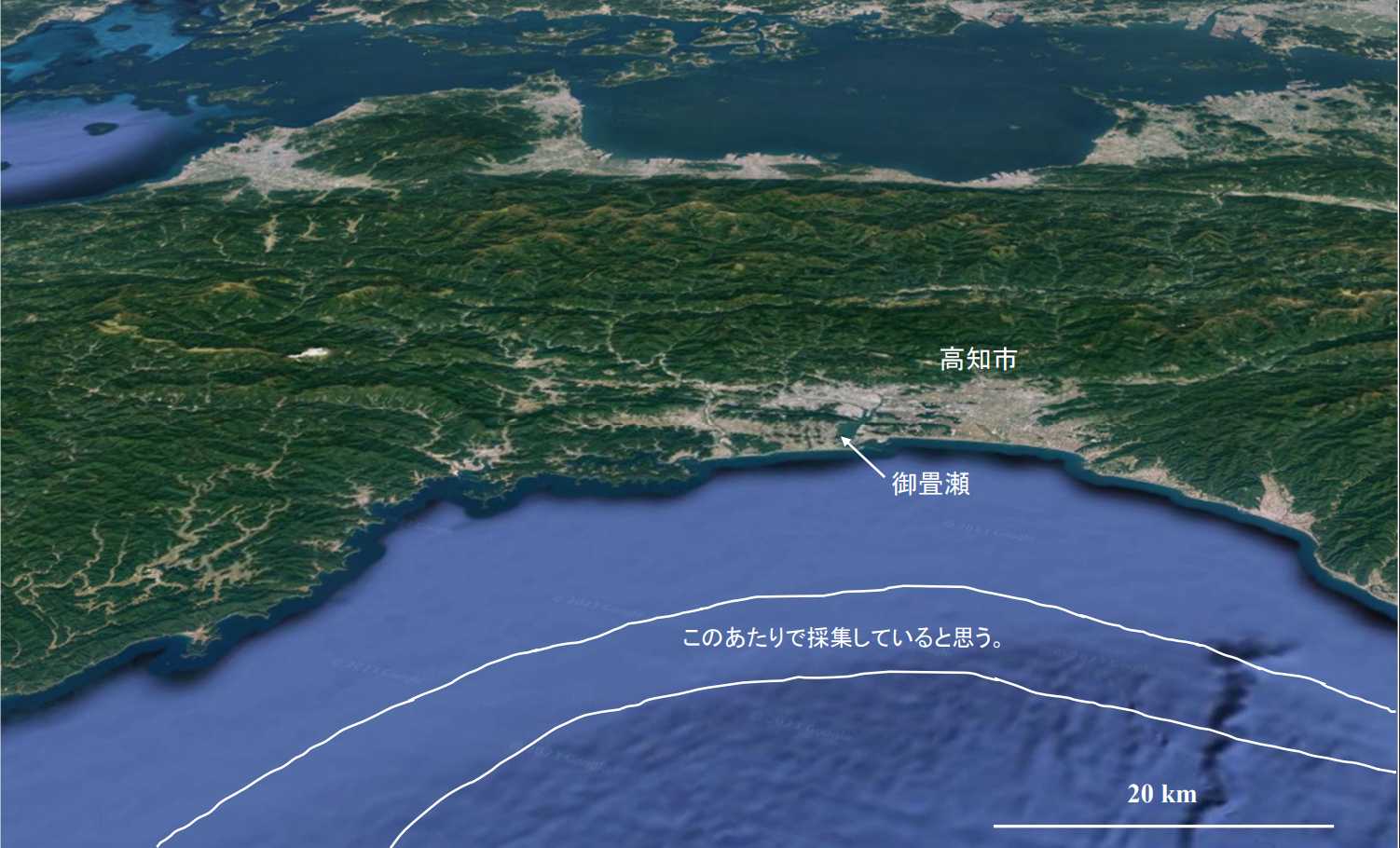
図 18.底引網を行う海域(深さ 150m~350m)。船(司丸)はいつ出港するか不明。夜出て明け方から操業をはじめ,ある程度の漁獲量が確保できたら,御畳瀬(みませ)に戻ってくるのではなかろうか?いつ頃戻るかも不明。早ければ 15 時ごろ,遅ければ夜遅くになることもあるだろう。底引網で捕獲する漁獲類は主には魚類で,メヒカリ(Chlorophthalmus albatrossis),レンコダイ?(Dentex tumifrons),サメ類など,いろいろな種類があった。メヒカリは干物に,レンコダイやサメはかまぼこの材料になるのだろう。アカザエビ,ウチワエビ,タカアシガニの他,根鰓亜目のエビ類が大量に入る。

図 19.オサテエビ(Thaumastocheles japonicus)。アカマダラアナエビからガラッと変わって.オサテエビを示した。土佐湾の深さ 200m~300m ぐらいの深海の底引網で採集できる。MP プレート境界はアカザエビと同じ「コ」の字型。中央の棘はあるが,アカザエビのように「根ほり」状の突起ではない。顎脚および歩脚の各プレートの融合が進んでいる。頭胸部の各プレートの融合がどの程度進んでいるかは,種類(グループ)によって大きく異なる。プレート間の溝の形成に関与する遺伝子は多分あるだろう。その遺伝子の機能異常がプレート間の融合につながるような気がする。一方,プレート間の溝の形成に関与する遺伝子によって,グループに特有な溝のパターンの形成に関するメカニズムの質問については,現在の段階では回答が難しい。

図 20.オサテエビ(Thaumastocheles japonicus)の右側の鋏脚(第 1 歩脚)の形態。オサテエビは,第 1 歩脚(いわゆるハサミになった付属肢)の構造が右と左では大きく異なる。右の鋏脚は長く,一列の棘が生えている。対面の掌節も長く,やはり 1 列の棘が生えている。右側の鋏脚で物(例えば,藻類の切れ端)をつかむのであろう。生息場所の水深は 200m から 300m ぐらいだろうから,水面からの光は全く届かず,完全暗黒の世界で生活していると思われる。右側の長い「箸」を使って恐る恐るエサを探し,食べられそうなものがあったら箸を使って手元に引き寄せるのだろう。深海では,とんでもなく長い鋏脚を持った十脚類が生活している。特にアンコウみたいな魚類は,体に触れる刺激があると,所かまわず,一気に噛みついてくるのだろう。

図 21.オサテエビ(Thaumastocheles japonicus)の左側の鋏脚(第 1 歩脚)の形態。オサテエビは,第 1 歩脚の形態が右と左では大きく異なる。左側の鋏脚は,多くの十脚類の鋏脚の形態によく似ている。左右の鋏脚で形態が大きく異なる種類としては,キバウミニナ(巻貝)を食べるオウギガニがいる。右側の長い鋏脚は貝の中に差し込んで肉(身)を切り刻み,左側の鋏は切り出した肉をつかんで口に持ってゆく。鋏脚のつかい方に機能分担がある。

図 22.アメリカン・ロブスター(Homarus americanus)。カナダ産(採集日・採集場所は不明)。ロブスターには,H. americanus と H. norvegicus の 2 種類がいる。いずれも形態は,淡水域に生息しているザリガニ類と非常によく似ている。ロブスターもザリガニも,やはり U 字型の MP プレート境界溝がある。ロブスターの祖先は,抱卵亜目(Pleocyemata)の最も古いグループに近いだろう。あるいは抱卵亜目の最初のグループなのかもしれない。

図 23.オキナワアナジャコ(Thalassina anomala)。英語名称は mangrove lobster。MP プレート境界線は,ロブスターと同じ「つぶれ U 字」型。ロブスターに比べると MP プレート境界溝の後半の部分が(相対的に)肥大している。頭部の棘(rostrum)は小さくなっている。化石は新生代から出ているが,中生代には祖先はどこでどんな姿で生活していたのだろうか? 祖先はロブスター系だろう。2017 年 4 月 21 日,西表島古見のマングローブで採集。
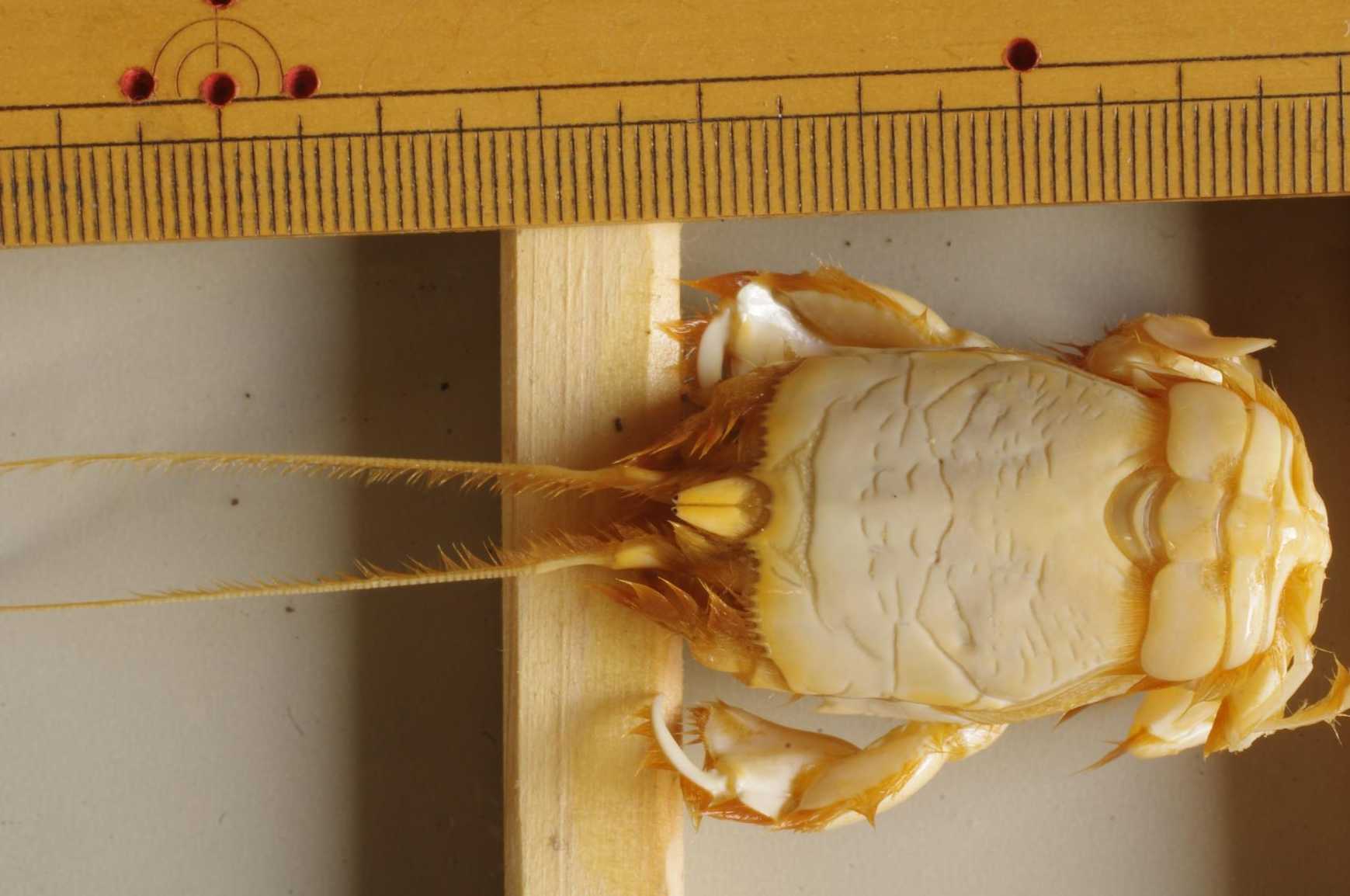
図 24.クダヒゲガニ(Albunea symnista)。MP プレート境界溝は,つぶれ M 字型。眼は,長い間の暗黒生活で退化していると思うが,黒い色素があるので,明暗ぐらいは識別できていると思われる。2004 年 6 月 4 日,西表島の潮間帯で採集。船浦か干立西側の潮間帯で採集した。クダヒゲガニは,土佐湾の比較的浅い潮下帯でも,イセエビの網にかかる。太平洋の広範囲の浅海に分布していると思われる。

図 25.クダヒゲガニの腹側。歩脚の配置のパターンは,ヤドカリ上科とホンヤドカリ上科と同じ。もうひとつ重要な特徴は,第 3 顎脚の付け根の後ろにある小さな隔壁(矢印)である。この突起は,ヤドカリ上科には見られず,ホンヤドカリ上科に共通する特徴である。ヤドカリ上科とホンヤドカリ上科は,三畳紀後期の Gastrodoridae(化石種)から進化したとされる。ヤドカリ上科とホンヤドカリ上科が別れた後に,ホンヤドカリ上科の中から浅海の砂浜に潜るグループが現れ,それが現在のクダヒゲガニになったと考えられる。ホンヤドカリ上科から分かれたのはジュラ紀のことだろう。現在では分岐年代の推定が可能なので,その結果(BEAST2)を加えれば,原著論文としてパブリッシュできるだろう。

図 26.クダヒゲガニの生息する砂浜海岸(西表島,船浦)。2004 年 5 月 27 日。中央やや左側に見えるのはヒナイ大橋。正面は琉球大学熱帯圏研究センター,右側の方は船浦の集落。この辺一帯は遠浅になっていて,潮が引いているときに浅瀬(水かさ 10 cm 以内)を歩くと,クダヒゲガニが飛び出すことがある。水の中を勢いよく泳いでから砂の中に隠れるので,隠れた場所の砂をすくえば採集することができる。ただし,効率は悪い。刺し網でも置いておけば,少しはかかるかもしれない。一番良いのは,何度も訪れてその都度お宝を少しずついただいて帰るということだろう。何度も・・・です。
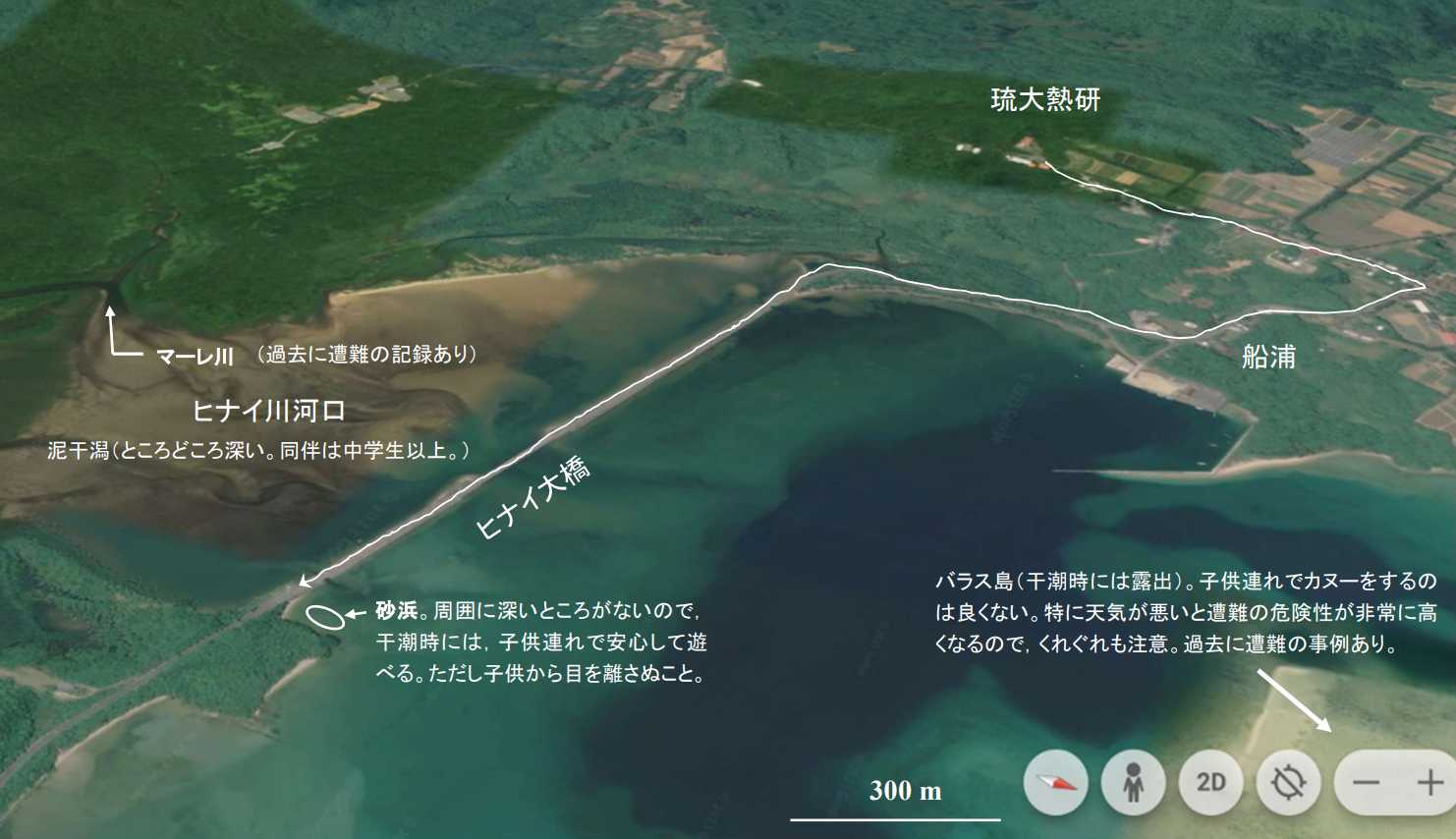
図 27.船浦湾の地形。ヒナイ川河口には,干潮時になると広大な泥干潟が現れる。泥干潟の上にはミナミコメツキガニの大群が現れる。また,砂地の多種硬いところにはユムシが触手(?)を出している。砂地の上にはヒメシオマネキが多い。周辺のヒルギ林(マングローブ)には,オキナワアナジャコの巣穴(本体を見ることは稀),多くのイワガニ類,シレナシジミ,ミナミトビハゼ等を見ることができる。ヒナイ川の河口は,ヒナイ大橋ができてから生物相は大きく変わったと思われるが,泥干潟は割と安全な場所である。ヒナイ大橋の外側でも,底質が砂地のところが多く,干潮時には子供連れで遊べる。一方,西表島は天気が崩れると,川はすぐに増水して氾濫を起こす。どこで崖崩れが発生するかわからない。海では激しい突風が吹き,急に強い波が押し寄せ,カヌーは転覆する。天気が崩れたときには外出するべきではない。余裕をもって自然を楽しむ,という心構えが必要である。