
令和5年(2023)1月29日(日)
近澤峰男さんは,お亡くなりになる前に,ご自身が撮影された野鳥と風景の写真ファイルを送ってくださった。その意図は,ご自分が撮影された多くの写真を,私の研究と教育に役立ててほしいという願いだったと思う。
近澤さんの撮影された写真を私の研究や教育に利用する大きなメリットは,質の高い写真を使えば,私の行う記述(description)の説得力が著しく上昇し,研究と教育内容の顕著なレベルアップが期待できる点にある。そんなことはまやかしだと非難する方々は多いだろう。
しかし,写真には,確かに文字(word)だけでは難しい生命現象を容易に表現できる大きなメリットがある。問題は,写真だけ提示したのでは,これまた何を言おうとしているかよくわからない点にある。写真と文字のコラボレーションが,現代科学を推進する強力な武器になる。私がやるべき事業(非営利活動)は,近澤さんが撮影された写真(No. 4Bの中にはない)や自分自身で撮影した写真を使って,大学・研究所・博物館・各種の学校・各地の自然保護センターの行う様々な学習活動に利用できそうなテキストを作ることだろう。
もうひとつ。多くの写真家の提示する画像や科学者の書く本には,「君たちも自分たち(写真家や科学者)をまねて,熱心に写真を撮り続け,熱心に研究を続ければ,自分のように立派な人物になれる。」という親切なアドバイス(要するにバイアス)が各所に感じられる。近澤さんの写真には,そういうバイアスが全く感じられない。だからこそ,近澤さんの写真は,私の研究と教育に大いに利用できるのだと思う。

図 1.サンゴ礁原における生物の撮影。サンゴ塊の下や隙間にやみくもに手を突っ込むのは危険である。
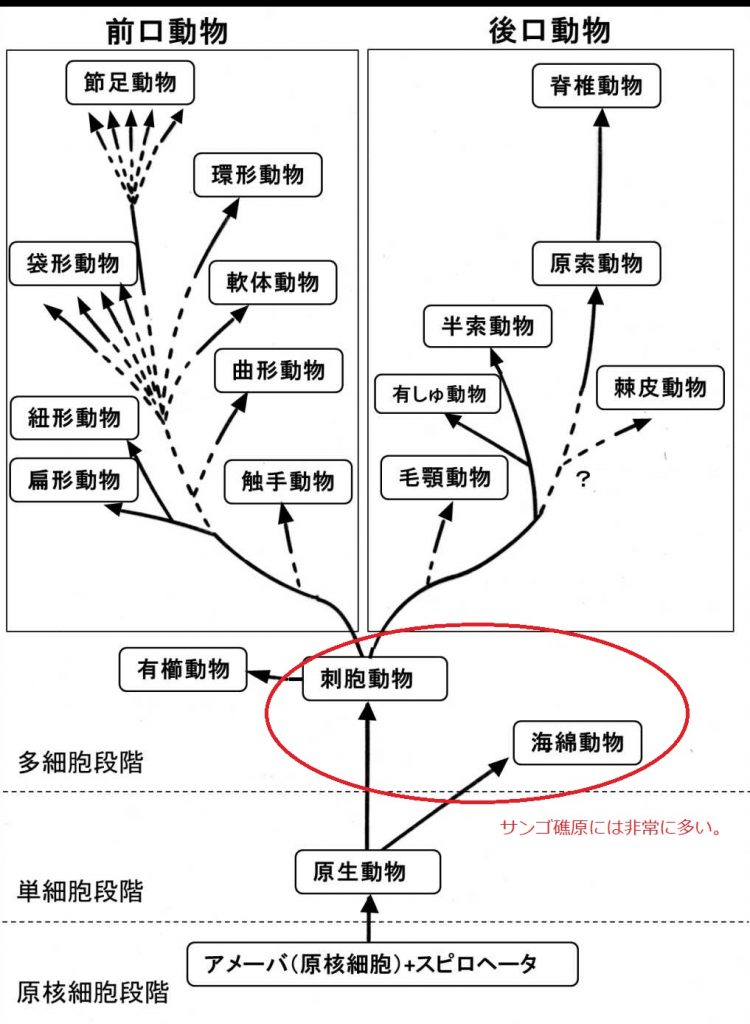
図 2.動物の系統図(原図)。現在はどこの教科書を見ても,同じ系統図が使われている。
多細胞動物は,原生生物が細胞分裂を起こした後に互いに離れず,接着したことがきっかけだったと思われる。では,多細胞生物の組織や器官はどのようにしてできたか?
私の授業内容は,高等学校の生物の教科書と大きな齟齬(そご)はないと思う。これは高校生物の教科書に遠慮したからではなく,教科書を作る際に参考にした文献が同じことに起因する。しかし,違うところも多くある。私の授業では,高校生物の記述とどこが違うか(主には生物現象に対する見方)を指摘することは多々あったと思う。もちろん,違うからと言ってマスコミを味方につけて訴え出ようなどとは,決して思わない。根拠が希薄な中で,どこが違うかという話は生産的ではない。そういう話は置いといて,高校の生物の教科書の延長として,海洋環境の成り立ちと,そこにはどんな生物がいるかを,それぞれの分類群(門)の系統関係を考慮して紹介したい。
さて,どの教科書を見ても,地球上に生命が誕生したのは,46億年前と書かれている。最初の生命が誕生した過程は,オパーリンが提唱したコアセルベート説がある。古い説であるが,説得力はあると思う。最初の生命は,原核生物(procaryote)の細菌(bacteria)だったのだろう。バクテリアの世界は,いわゆる細菌(bacteria)と光合成をする細菌(日本では原核性の「ラン藻」ということが多い。)の世界は,何と20億年も続いた。
先カンブリア代の15億年前あたりから,生物の構造に変化が現れた。つまり,原核細胞の中に寄生する単細胞が現れ始めたことである。寄生(parasitism)は生物の持つ基本的な性質なので,原核細胞同士で起こっても不思議はない。時間はかかったが,寄生の結果として,菌類(fungi)(細菌ではなく,カビやキノコの類),植物細胞,そして動物細胞などが出現した。この時点ではすべて単細胞生物(Whittakerの言う原生生物)である(図2)。アメーバ,ゾウリムシ,ミドリムシなど・・・(図はない)。

図 3.赤離(アカパナリ)の潮間帯。大潮の干潮時であるが,潮(tide)がかなり引いてきたところである。サンゴ礁原に行くには,潮汐表(インターネットで見れる)を調べて,2 時間ぐらい前に海岸に到着するのがよい。なお,荷物は西表島では,車の中に置こうが,岩の上に置こうが構わないが,外国,例えばグアムでは窃盗が多い。ついてすぐ,臨海実験所(Guam Marine Laboratory)の所長から,荷物を車に置く場合にはトランクに入れ,カギをかけるのを忘れないようにとの注意を受けた。何と,私たちが来る直前にいろいろな家具をそろえたが,テレビを除いてみんな盗まれてしまったとのこと。
動物細胞(図2)の最も重要な特徴は,細胞の表面に鞭毛(べんもう)や繊毛(せんもう)が生えていることである。べん毛と繊毛は透過型電子顕微鏡(Transmission electron microscope)で断面を見ると,基本的な構造は同じなので。両者の起源は同じと推定される。ゾウリムシ(繊毛虫類)とミドリムシ(べん毛虫類)は,基本的には同じ原生生物と考えてよい。
動物は原生生物(Phylum Protozoa)の中から進化した。原生生物の中にはミドリムシ(Euglena)含まれる。Euglenaは,細胞内に葉緑体を持っているので,動物はEuglenaから進化したという可能性は非常に低いだろう。
動物は脊椎動物(vertebrates)と無脊椎動物(invertebrates)の2つに大別される(図2)。それらの分岐点に位置するのが刺胞動物(Phylum Cnidaria)である。それから前口動物(Protostomia)と後口動物(Deuterostomia)に分かれた。なお,高校の教科書では,旧口動物と新口動物という呼称がずっと続いている。
前口動物と後口動物の大きな違いは,胚発生(embryonic development)にある。詳しいところは見ていないが,原口(blastopore)(胞胚において原腸胚形成中に形成される陥入部の入口)が口になり(つまり先に口ができて),あとから肛門ができるのが前口動物(旧口動物),原口の陥入部が肛門になり,後で口ができるグループが後口動物(新口動物)である。「前」というのは,「先にできる」という意味。「後」というのは「後でできる」という意味である。
Wikipediaを見ると,「扁形動物・刺胞動物・有櫛動物(図2)では消化管は途中で行き止まりになっている。従って,消化管の出入り口は1つしかない。これが原口に相当すると考えられる。要するに,扁形動物・刺胞動物・有櫛動物は新口動物でも旧口動物でもない。」と書かれている。図2(私が描いた)では,扁形動物(プラナリアなど)は,前口(旧口)動物になっている。
原生動物(Phylum Protozoa)は,すべての種類がいつまでも単細胞でいた訳ではない。一部の原生動物の中に細胞分裂をした後に,互いに細胞が分離せずに,接着した状態の細胞塊が出現したのだろう。
多細胞化して最初に現れた生物は,動物では海綿動物(Phylum Porifera),植物では藻類(algae)だろう。海綿動物は個体性が低く,多細胞の藻類(例えば緑藻類のアオサ)とよく似た体の仕組みになっていただろう。植物では,緑藻類から維管束植物が進化したが,動物では海綿動物から進化した高等な生物はいないとされる。
海岸に出てみよう(図3~図6)。サンゴ礁原(潮間帯)では,潮が引かないと海産動物の採集は難しい。私たちが行くのは,いつも大潮(spring tide)の干潮時(low tide)である。大潮の時期は,満月や新月の頃の5~6日間である。大潮の時期は,潮汐(ちょうせき)現象に関わる地球・月・太陽が一直線上に位置する。そのため,月と太陽の持つ起潮力が加算され,1日2回あるうちの干潮(low tide)時には,潮は上弦や下弦の頃よりずっとよく引いている。特に太平洋岸は,3月から5月の潮は,夜間より昼間の方がよく引くため (つまり潮間帯の最も下部まで引くということ),普段だと潜らなければ見られない多くの海産生物を目の当たりにすることができる。
ただし,3月から5月は,日差しが非常に強いので,Yシャツを着ていても,激しい日焼けを起こすので注意されたい。日焼け止めのクリームは必需品である。

図 4.大潮の干潮時のサンゴ礁原。昼間にこんなに潮が引くのは,3 月から 5 月いっぱいである。

図 5.サンゴ礁原に現れたタイドプール。

図 6.タイドプールで泳ぐ魚たち。魚を追いかけると無脊椎動物を観察する時間が無くなる。

図 7.サンゴ礁原での無脊椎動物の撮影。撮影機材は,Optio(PENTAX)。水中でも撮影可能。
図 4 は,潮が完全に引いた時のサンゴ礁原を示している。場所は赤離(アカパナリ)だろう。サンゴ礁原と言っても,水面から露出する範囲(図 4 と図 7)では,生きているサンゴ(塊)は少なく,死んだサンゴ塊の上におびただしい数の無脊椎動物(invertebrates)が,所狭しと体を寄せ合って群体を作っている。タイドプールの中には生きているサンゴ塊がみられる(図 5と図6)。図7は干潟で生物を撮影していところ。これだけ干出すると生きたサンゴはいない。

図8.サンゴ礁原のタイドプールの中で見かける典型的な生物群集。固着性の生物は,サンゴ(刺胞動物),イソギンチャク(刺胞動物),カイメン(海綿動物)が多い。
西表島のサンゴ礁原(干潟)に見られるタイドプールの中は,図8のような景観になっている。この中に何種類ぐらい生物がいるのか,実はよくわからないが,相当多いだろう。
真ん中と左側のピンク色は,イソギンチャクのポリプかもしれない。その間にある小豆を包んだ白いアイスクリームのような動物は,やはりサンゴのような気がする。写真(図8)左下の小豆を包んだあずき色の動物もサンゴ,いわゆるソフトコーラルかもしれない。その上の黒いすすみたは,カイメンだろう。カイメンの付近は不明だが,サンゴの可能性が高い。
右側の黄緑色の突起は,サンゴの一種だろうか。中央下側に見える紅色の動物も,やはりサンゴだろうか。私の知識不足もあるだろうが,タイドプールの中には門(Phylum)の名称すら不明なたくさんの無脊椎動物が生息している。門すらわからないのだから,その下部の分類群(綱や目)に至っては全く不明である。
サンゴ礁原の代表的な海産生物の中で,割とわかりやすいのはカイメン(海綿動物門: Phylum Porifera)である。
カイメンは英語ではsponge。確かにスポンジみたいな雰囲気はある(図9~図11)。

図 9.サンゴ塊(死骸)の表面に繁茂するカイメン。綱(class)の分類は,私にはよくわからない。

図 10.生きたサンゴ塊の上に広がるカイメン(種名はわからない)。カイメンの下には生きたサンゴがある。

図 11.サンゴ塊(死骸)の表面を覆っているカイメン。中にテッポウエビ(十脚甲殻類)が入っている。テッポウエビの右側の鋏脚(かんきゃく)が見える。写真の左側には小さい巻貝(軟体動物)が 7 つほど見える(5 種類ぐらいか?)。巻貝(これも種類は不明)は,サンゴ塊(死骸)の上に広がったカイメンの上にいる。
カイメン動物(Phylum Porifera)は,以下の4つの綱(Class)で構成されている。4つの綱が多いと言えるのか,多くないと言えるのか,私にはよくわからない。
<カイメン動物門>
1)石灰海綿綱 Class Calcarea
2)普通海綿綱 Class Demospongiae
3)六放海綿綱 Class Hexactinellida
4)硬骨海綿綱 Class Sclerospongiae
カイメンは,体表に数多くの小孔を持ち,小孔を通して海水の循環を起こしている。体の内側は空洞(胃腔)になっていて,鞭毛をもった襟(えり)細胞が多数並んでいる。海水は,襟細胞から出ている鞭毛の動きによって胃腔内で循環している。食物(海水中の懸濁物)は,小孔や胃腔内の表面にある襟細胞のべん毛によって捕獲され,襟細胞によって細胞内消化され,栄養としてカイメンの成長に利用される。胃腔内に集まった海水は,体の上部にある大孔から吐き出される。
なお,胃腔という漢字は,「いこう」と読むか,「いくう」と読むか,よく知らない。生物学の分野では,腔腸(こうちょう)動物と言うので「いこう」と読むかもしれない。
医学系の分野では口腔(こうくう)外科だから「いくう」と読むことになる。もっとも,医学系の分野でカイメン動物を実験材料として使うことはまずないのだが・・・。
同じように,神経細胞(neuron)から延びる軸索(axon)については,生物学分野では「神経繊維」と書くが,医学系の分野では「神経線維」と書く。こっちの方は,高校生物の教科書を編集するときに問題が生じる。
私は,教科書の執筆者によって違った書き方があっていいと思う。こんな話も授業の中に含めたら,関心を持つ学生もいるのではないか?
カイメン動物は,系統進化学的には側生動物(Parazoa)と言われている。つまり,カイメン動物は,多細胞になっているが,組織や器官が分化(differentiate)していない(図9~図11)。刺胞動物のように神経細胞が存在しないので,多細胞体として統一の取れた動きができない。個体性(individuality)の程度は低いと考えてよい。菌類(fungi)で言えば,カビやキノコと同様な体制とみなしてよいだろう。
もうひとつ。カイメンは,多細胞動物の中でも最も下等な体制(lowest organization)を持ち,群体を作る。Googleで「体制の最も下等な」で検索すると「most inferior system」と出てくるが,この訳はいただけない。人間の価値観で動物を比較するのはまずい。辞書を信用してこんな表現を書くと,書いた人の見識が疑われるので十分に注意していただきたい。
「群体」を日本大百科全書(ニッポニカ)で検索すると「分裂または出芽によって生じた新たな個体が互いに連結してできた個体の集合である。原生動物・海綿動物・刺胞動物・コケムシ類・ホヤ類(原索動物)など,海産無脊椎動物の多くのグループにみられる。群体を形成する個体を個虫と言う」とある。
ニッポニカさん,ちょっと待ってくれ。単細胞と多細胞における群体をいっしょにしないでくれ。群体を形成するのは,原生生物(ゾウリムシ)やカイメン動物では,細胞である。刺胞動物・コケムシ・ホヤでは,細胞ではなく多細胞で組織や器官のある個体である。いっしょにしてはまずい。
このあたりのことになるとついて来られる学生の数は著しく減少し,スマホをいじっているか,寝ているかのどちらかになる。アニマル浜口氏ではないが,大きな声で「重要だ!重要だ!重要だ!」と3回叫んで,スマホに目を向けないように注意喚起を行う。授業時間内に授業内容をまとめさせていれば,スマホいじりと寝ることはいくらか防げるかもしれないが,5分もすれば,多くの学生の目の動きはまた元の姿に戻る。しかし授業を聴いている学生もいる。
次の問題は,同じ群体と言っても,カイメンとイソギンチャクやサンゴでは少し意味が違うことである。カイメンの群体は,細胞の集合体で固有の形態が作られない(図9~図11)。一方,イソギンチャクやサンゴは,細胞の集合体が固有の形態(個虫)を 作り,さらに個虫が集まって群体を作っている。
この問題を議論する前に,カイメンと藻類の形づくりの共通性について述べておきたい。図12は,サンゴ礁原で見かけた藻類を示している。間違えたら申し訳ないが,動物ではなさそうだ。私には褐藻類の1種のように見える。これは群体ではないのだろうか?カイメン(図9~図11)とはどう違うのだろうか?
もう一つの例を示そう。図13は緑藻類の群体を示している。これも図12の褐藻類と同じく,多細胞生物(緑藻類の個虫)の群体だろう。これもカイメンとはどう違うのだろうか?
結論を言えば,図12の褐藻も,図13の緑藻も「群体」とは言わない。植物の場合には,習慣的に群体とは呼んでいないようである。しかし,生物の体制(organization)を見れば,図12の褐藻と図12の緑藻は,カイメンの体制(図9~図11)よりも少し高等になって,イソギンチャクの群体(図14)と同程度に達しているとみなすことはできないだろうか?
イソギンチャクには散在神経系(diffuse nervous system)が備わっていて,個体として統合的な動きが可能になっている。しかし,褐藻や緑藻を見れば,生物の固有の形(フォルムあるいはmorphology)は神経系がなくても生じうることがよく理解される。最初は偶然にせよ,できた形態は,自然選択のふるいにかけられたのだろう。自然選択のふるいによって,時間とともに個体性が高まったと考えられる。神経系は,出来上がった体の各部分(組織や器官)の動きを調整して,効率よく行動できるために使われているのだろう。

図 12.サンゴ塊の上に見られた褐藻類の個虫の集合体(群体)。個体性が確立している感じである。

図 13.緑藻類の個虫の集合体(群体)。個虫の個体性は確立している感じだ。

図 14.イソギンチャク(刺胞動物)の群体。個虫の個体性は,カイメン(図 8 から図 10)よりはるかに高く,図 11 と 12 に示した褐藻類や緑藻類と同じ段階に達しているように思われる。

図 15。イソギンチャク(種類は不明)の群体?あるいは触手を持った単体かもしれない。

図 16.これはイソギンチャクの群体のように見えるが,個虫が見当たらない。つまり図 14 のように個虫の個体性が確立していないので,カイメンの一種かもしれない。藻類ではなさそう・・・。すみません,何か全然わからない。およそ似つかわしくないが,カイメンの一種と思う。

図 17.イソギンチャク(種名は不明)。クマノミがいるので単体(一個体)のイソギンチャクとわかる。
カイメンの次は,サンゴ,クラゲ,イソギンチャクの含まれる刺胞動物(Phylum Cnidaria)である(図2)。刺胞動物の特徴は,カイメン動物と比較して「個体性」が高くなっていることだろう。図10と図14を比較すれば,その違いは一目瞭然である。
刺胞動物は,ヒドロ虫綱(カツオノエボシ,),箱虫綱(アンドンクラゲ),鉢虫綱(ミズクラゲ),花虫綱(多くのサンゴとイソギンチャク)などに大別される。
イソギンチャク(図14~図17)とサンゴ(図18)は,花虫綱(Class Anthozoa)に属する。Wikipediaを見ると,イソギンチャク(sea anemone)やサンゴ(coral)は,生活環のすべての時点でポリプ型の生活様式をとると書かれている。
サンゴ礁原の形成に関与するのは,主に花虫綱イシサンゴ目に属し,造礁サンゴ(reef-building corals)と呼ばれている。造礁サンゴは,サンゴ礁生態系の根幹をなす重要な生物である。造礁サンゴの面白い性質についてひとつだけ述べておきたい。

図 18.群体の成長によって生じる群体同士のギャップ。多細胞生物が組織や器官を作ることができたのは,このような性質があったからだろう。増殖の最初の頃は,群体に参加する個虫はみな「自分」になってくれるが,時間がたつと群体に参加する個虫は,遺伝子型が全く同じであっても,「他人」になってしまう。だから「他人」になった段階で,新しい組織や器官を「自分」とは独立して作ることができたのだろう。
サンゴは無性生殖(おそらく出芽)によって増殖する。増殖したサンゴは,多細胞で個虫となって隣り合わせる他の個虫に付着して群体を形成する。図18を見ると,海ブドウ(緑藻類)の周囲に群体が形成され,さらに成長してゆくと反対側から成長した群体と接することがある。群体を作るからには,隣り合う個虫は「自分」と同じ性質を持っているはずなのだが,群体が成長するに従い,他の側から成長してきた群体と接するところにギャップができる。ギャップができるということは,個虫が分裂してゆく間にだんだん「自分」とは違った性質を持つ個虫が育って行くことを示すのだろう。
カイメンや藻類は群体を形成するにあたって,群体に参加するそれぞれの細胞が,遺伝子発現を含めていつまでも「自分」と同じ性質を持つ。この点が,個虫が集まって群体を形成するサンゴ・イソギンチャク・クラゲ(刺胞動物)と大きく異なっている。刺胞動物では,個虫の分裂にともなって,だんだん「自分」から「他人」に変わる。このあたりに生物進化の根源的な原因がありそうだ。
<結論>
生物は,単細胞から構成される群体であれ,単細胞が集まってできた個体から構成される群体であれ,要素(単細胞や個体)の単なる集積では説明がつかない。水を含めて物質は,どんなに量が増えても,それぞれの分子の物理・化学的性質は変わらないが,生物は細胞が増えて行くと,最初に分裂したことの細胞と,後になって分裂した細胞とでは,性質が異なってくる。上でカイメンや藻類が分裂後もいつまでも「自分」と同じと書いたが,厳密には,分裂を繰り返すうちに細胞の性質は,少しずつ違ってくるのだろう。遺伝子発現に違いがあるかは不明。この性質の違いが組織・器官・個体の形成に大きな役割を果たしているのではないか?

図19.高那(たかな)のサンゴ礁原。イソギンチャクの巨大な群集。サンゴ礁原では,優占種は,ほんのわずかな地形の違いによって大きく異なるようだ。近いうちに生態系保全学の視点から,サンゴ礁原の形成過程について説明してみたい。
図19は,西表島の北側に広がるサンゴ礁原である。タイドプールの中には生きたサンゴがみられるが,ここはイソギンチャクがびっしりとサンゴ塊(死骸)に張り付いている。(イソギンチャクの種名はわからない。)
座礁したのは鋼材輸送船で,名前は鉄栄丸と言うらしい。鉄栄丸は,戦後何か所かで複数隻が建造されているようだ。写真に写っているのがどの鉄栄丸かよくわからない。(インターネットの記事は信ぴょう性に欠ける。)アカパナリのサンゴ礁原の形(地形)は,船浦のそれによく似ている。船は船浦港と間違えてサンゴ礁原に乗り上げたと思われる。1970年代の頃だろう。目視で航行していたと思われる。あと50年したら海の中に消えてしまうかもしれない。
難破船の話は置いといて,石垣島や西表島では,サンゴ礁原の景観は,島の東側と西側では大きく異なっている。
サンゴ礁原の生物はほんのわずかな環境条件の違いに左右されて成長と消滅を繰り返している。そのため,礁原(潮間帯)の景観は島によって大きく異なってくる。次は,生態系保全学の視点から見た宮古島のサンゴ礁原(潮間帯)について述べてみたい。
<参考文献>
- 吉野治一(2015)水に学ぶ物質科学。http://e.sci.osaka-cu.ac.jp/yoshino/edu/water/water.pdf
- Ehsan, K., B. Bastian, S.M. Pankey, A. H. Ohdera, M. Monica, P.C. David, A.G. Collins, and R.F. Ryan (2018) Phylogenomics provides a robust topology of the major cnidarian lineages and insights on the origins of key organismal traits. BMC Evol. Biol. 18: 68. doi:10.1186/s12862-018-1142-0.
- Fieseler, L., M. Horn, M. Wagner, and U. Hentschel (2004)Discovery of the novel candidate phylum “Poribacteria” in Marine Sponges. Appl. Environ. Microbiol. 70: 3724–3732. doi:10.1128/AEM.70.6.3724-3732.2004.
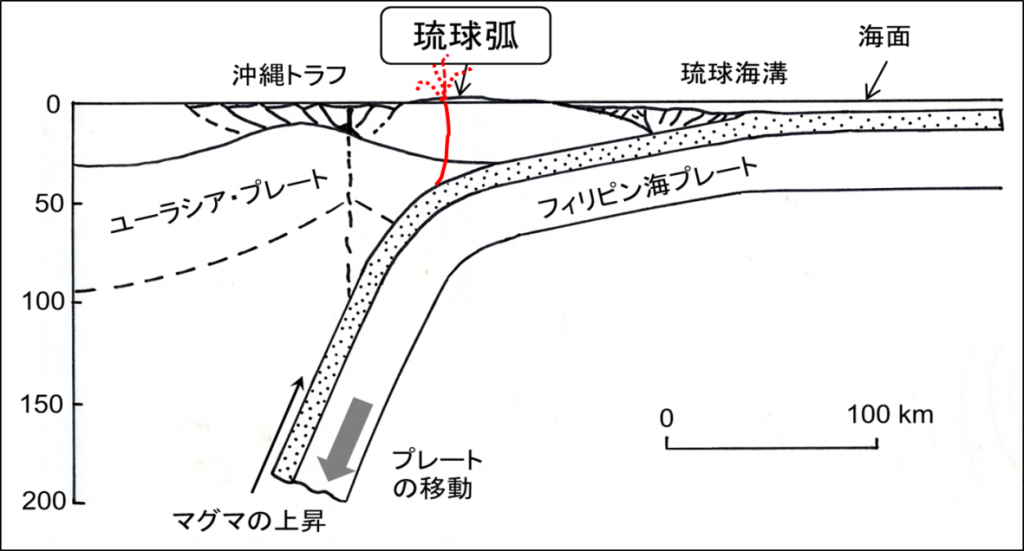
<資料>琉球弧の成因。九州からトカラ列島にかけては,マグマの噴出で,休火山を含めて火山島が多くみられる。奄美大島から八重山諸島にかけては,マグマの噴出はなく(琉球トラフではあるかもしれない),島々は堆積岩や琉球石灰岩でできている。