
2024年10月5日(土)
アダンの育つサンゴ礁の島
鳩間島はヤシガニ(Birgus latro)で有名だが,実際にどんな環境にすんでいるのか,また生息個体数はどれほどかの情報はなかった。今年(2024)の6月30日に鳩間島を訪れ,アダンが島の植生の大半を占めること,そしてヤシガニはアダンの林の中に隠れていることを知った。ヤシガニは,ベンケイガニ類のように土手に巣穴を掘って棲んでいる訳ではなかった。
鳩間島に生息しているヤシガニの個体数がどれぐらいか不明であるが,直感で推定すると島全体で百匹少々というところだろうか。乱獲すればすぐに絶滅する数である。ヤシガニは,インド洋から太平洋のサンゴ礁に囲まれた多くの島々に生息する。その意味では普通種(common species)と言える。しかし,十脚甲殻類(decapod crustaceans)の中では,タカアシガニ,ズワイガニ,キングクラブ(オウギガニ?)と並んで,1個体の大きさは半端なく大きくなる。ヤシガニの生息域は,島の大きさに関わらずサンゴ礁の海岸沿いの陸上に限られる。だから,どこの島でも生息個体数は少なくなるのだろう。要するに,ヤシガニはどこに行っても希少生物の仲間に入る。
このような事情から,鳩間島に生息するヤシガニは保護する必要がある。捕獲禁止は,保護のひとつの手段として有効である。また捕獲禁止だけでなく,ヤシガニの生活場所であるアダンの林も同時に守ってやるとよいだろう。
面白いことに,アダンは昆虫類の食害をほとんど受けない。何か昆虫の忌避物質を含んでいるのかもしれない。忌避物質が精製できれば,有害昆虫に対する防除に利用できるかもしれない。アロエと見かけが(ちょっとだけ)似ているので,漢方薬を作れる可能性もあるが,有毒な物質という可能性の方が大きそうだ。
アダンは,熱帯域の海岸や河口域では優占種(純林)となり,分厚い層の密林を形成する。ヤシ類とは大違いである。赤道付近にあるサンゴ礁の島々では,あの凶悪なアダンの密林では,マラリアを媒介するハマダラカがそれこそ雲のごとく襲ってくる。太平洋戦争で南方の島々に派遣された兵隊が,ハマダラカの大群の襲撃を受けながら,山刀を片手にアダンの密林(湿地帯)を切り開いて進んだのだろう。やっと切り開いた密林の先には,アメリカ軍の集中砲火が待っていたのかと思うと胸が痛くなる。
西表島ぐらいの緯度(24.3°E)になるとアダンの密林の凶悪性は低下し,平地であれば,小さなブルドーザーがあればすぐに切り開けるところが多い。

図 1.仲間川(西表島)の土手にできたアダンの林。
熱帯から亜熱帯にある多くの河川では,満潮時に水(海水からほぼ淡水まで)に浸かる河口域には,ヒルギの群落ができる。水に浸からない土手にはアダンの林が形成される。この場所は海から 4 km ほど上流にあり,川沿いには淡水域に分布を広げているオヒルギの純林が見られる。アダンの背の高さは 1m ほどで,規模は小さい。それでも,アダンの生えている土手を登って熱帯林の中に入ってゆくのは難しい。細長く硬い葉の上に強烈なトゲが並んでいて,これが皮膚に刺さる。赤道付近の島々だと,アダンの背丈は 5~6m に達する。林の中はハマダラカの格好の住みかとなる。相当ぶった切ってからでないと土手を登れない。

図 2.船浦(西表島)の海岸にあるアダンの群落。
アダンは海岸沿いの砂泥岩層の上によく見られる。この場所では,砂泥岩が頻繁に崩れ落ちている。海岸に落ちた砂泥岩は,海に向かって少しずつ移動する。波に洗われるようになってから,50 年もすればイシアナジャコが巣穴を作って住みつくのだろう。土手は石や岩が崩れ落ちるので登ることはできない。さらに砂泥岩に表面にはアダンの群落がへばりついているので,とても上には登れない。土手の上にはリュウキュウマツ(裸子植物)やモクマオウ(被子植物)が見られるが,100 年以内にがけ崩れと一緒に倒れてしまうだろう。アダンの群落付近にヤシガニはいるかも知れないが,数は非常に少ないと思われる。(一匹いるかどうか?)海岸沿いのアダンの林にはオカヤドカリは多い。個体群密度の違いは,それぞれの種の個体の大きさ(size)を直接反映すると言い切ってよいのだろうか?
ヤシガニ:一属一種のオカヤドカリ
比較系統学の視点で見ると,ヤシガニの進化には現代生物学のアプローチを必要とする重要な課題があることがわかる。
まずは種類数。ヤシガニは,インド洋と太平洋の多くの島々に広く分布しているのに,1 種類しかいないのはなぜか?形態の微小な違いで無理に分けることはできるが,地域個体群の間で十分に交雑が可能だろう。だから,インド洋から太平洋の島々に分布するヤシガニは 1 種類と断定してよいのではないだろうか?ヤシガニと同様に,オキナワアナジャコも 1 種類(1 属 1 種)である。ヤハズアナエビやトゲスナモグリなども 1 属 1 種の可能性が高い。
ヤシガニの近縁種としてオカヤドカリ(land hermit crabs)がいる。オカヤドカリの方はヤドカリに比べて体の大きさは小さく,琉球弧には確か 5 種類ぐらいが分布していると思う。いずれも天然記念物に指定されているので,採集して研究に使用するには捕獲許可が必要である。
それは別な機会に議論するとして,海産動物の場合には,体が小さいと種分化(differentiation)が起きやすく,大きくなるに従って 1 種類に近づくというようなことは言えないのだろうか?
この原則は,幼生(larva)の生存期間と密接に関係する可能性がある。つまり,体が大きくなる種類(無脊椎動物)は,幼生期が長いのかもしれない。幼生期が長ければ,海流に乗って熱帯域の島々に広域に分布することが可能だろう。海を隔てて遠く離れた島々では,遺伝的には多少の違いは生じても,生殖隔離が起きる可能性は少ないかもしれない。

図 3.アダンの実。パイナップルとはだいぶ違う。鳩間島のヤシガニ生息地では,アダンの実は見つからなかった。ヤシガニが木に登って食べてしまったのだろう。なぜアダンガニと言わず,ヤシガニなのか?赤道周辺にある多くのサンゴ礁の島々では,海岸沿いには崖が少なく,そのような場所ではココヤシが優占種になっている。アダンの群落は海岸の崖や河口域の土手に形成される。そういう所には人はあまり行かないので,イメージとしては赤道周辺の島々では「ヤシガニ」となるだろう。

図 4.鳩間島の地形と自然環境。
西表島に初めて行ったのは,1971 年 3 月 12 日だったと思う。それから何十回も訪れていたが,上原からは目と鼻の先にありながら(高速船で 20 分)一度も行ったことはなかった。最近は上原から日帰りで往復できるようになったので,2024 年 6 月 30日に初めて訪れた。午前中は島の西半分を,午後には東半分を歩いて回った。鳩間島は宮古島と同じく,サンゴ礁原が隆起してできたのだろう。宮古島が海上に現れたのが今から 200 万年前ということなので,宮古島と同じ時期にできたのかもしれない。島の地層は,宮古島と同様に全域が琉球石灰岩(サンゴ礁が起源)でできていて,表面は琉球石灰岩が風化してできた赤土の層(薄い)で覆われている。

図 5.集落の外れから島の北に延びる小道。
集落内では樹木が多いが,集落を外れたとたんに景観が大きく変わる。道の両側には 2~3m の背丈のアダンがびっしり生えている。アダンの林は島の北側にある海岸まで続いている。埃道になってから廃棄物の置き場までは軽トラが通った跡がくっきりと残っているが,それより先は道の上に草が茂っている。ここから 10 分も歩けば北側の海岸に出る。

図 6.アダンの林。
アダンの林まで来た時に,林の中で何やらガサガサと動物の動く音が聞こえた。ヤシガニの生息地であることはすぐにわかった。アダンの林内は日中でも薄暗く,枯葉の下には隠れ場所も多い。ヤシガニにとっては良い場所かも知れないが,人間が入って行くには大きな抵抗がある。(5m も行けば方向がわからなくなるし,アダンの幹や枝が体に引っかかってなかなか前に進めない。)ブルドーザーがあればすぐに整地できるが,それではヤシガニの生息地を破壊することになる。
アダンの林は,放置すると 2~3 年もすれば道路に張り出して人や車が通れなくなる。・・・なので,定期的に林縁部を伐採する必要がある。誰がやるのかは知らない。詳細は調べていないが,ヤシガニは,竹富町自然環境保護条例で現在は 1 年を通じて捕獲が禁止されていると思われる。鳩間島にも条例が適用されるだろう。これだけ規模の大きい生息地は,琉球弧の中では鳩間島だけである。
次のステップとしては,ヤシガニがなぜ保護に値するかの理解を広めて行くことだろう。希少生物だからという理由では,納得しない人たちも多いのではないだろうか?だから法令で捕獲禁止を定めても,すぐに保護できるかは疑問である。保護を進めるための大義があるとよい。自然科学に限らず,研究は大義を見つけるために大いに役立つだろう。

図 7.道に出てきた中型サイズのヤシガニ。
体の各器官と歩脚の名称を示した。ヤシガニは,オカヤドカリが貝殻を抜け出して生活しているうちに,腹部の外骨格が再び分厚くなったのだろう。赤道付近のヤシが生い茂る小さな島々でオカヤドカリから分化したと思われる。その島にはよほど捕食者の数が少なく,またヤシの落ち葉で体を隠すことができたのだろう。最初はオカヤドカリほどのサイズだったが,世代を繰り返すうちに大型化したのだろう。ヤシガニの祖先がいつ頃貝殻を抜け出したのか,化石の記録からは判断しにくいと思われる。直感で言えば,オカヤドカリが貝殻を抜けたのが白亜紀で,体の方は新生代に入ってから大型化したような感じである。いずれ確実な証拠(化石がベストだが・・・)を見つけて再度記事にしたい。

図 8.ヤシガニ鋏脚(かんきゃく)の左右性(laterality)。
ヤシガニを含めてオカヤドカリの大きな特徴は,左右の鋏脚(かんきゃく)に機能分化が生じていることである。機能分化は当然形態の違いをともなっている。ヤドカリ類には左右の鋏の大きさが等しい種類,右が大きい種類,左が大きい種類がいる。ヤシガニを含め,オカヤドカリの仲間はすべて左鋏脚が利き手になっている。利き手の方(左鋏脚)で硬いもの(アダンの実など)を割り,もう一方(右鋏脚)でエサを挟んで口に運ぶ。
生態学者と生理学者(分子生物学者)の不協和音:十脚甲殻類における鋏脚左右性の発現をめぐって
十脚甲殻類では,利き手が失われると,1回目か2回目の脱皮の時に利き手の鋏脚が再生(regenerate)される。オカヤドカリでは利き手(左鋏脚)が失われても,脱皮の際に以前と同じように再生した左鋏脚が利き手になる。つまり,オカヤドカリの場合には,右を失おうと左を失おうと,再生した鋏脚の左右性は変化しない。また,潮間帯には右利きのヤドカリ(ホンヤドカリ)がいる。ホンヤドカリの右鋏脚を落としてみると(自切),右鋏脚は利き手として再生する。右利きのヤドカリの場合には,右鋏脚を失っても再び右鋏脚が利き手になる。
だから,右利きのヤドカリの場合には,右鋏脚が利き手になることに大きなメリットがあり,左利きのヤドカリの場合には左鋏脚が利き手になることに大きなメリットがあると考えることができる。
一方,カニ類は右の鋏脚が利き手になるが,右が失われた場合には,利き手は左鋏脚に移る。再生した右鋏脚は利き手ではなくなる。つまり,カニ類では利き手の喪失によって左右性が逆転する。もちろん,左鋏脚に移った利き手が失われれば,今度は右鋏脚が利き手になる。
ヤドカリやカニを使って行われた私たちの研究(Masunari et al., 2014, 2020)は,右利き・左利きに特別なメリットがある訳ではなく,利き手があることに大きなメリットがあることを強く示唆している。カニ類では,利き手が右から左に移るのは,右手が失われた結果であって,もともと左利きのカニがいる訳では無い。
生態学者は,右利きには右を使用する特別なメリット(例えば,右を使う方がエサを確保するのに都合がよいとか・・・)があって,左利きには左を使用する特別なメリットがあると考えている。・・・ならば,右の利き手が失われたとき,左右性が逆転するのはおかしいと思うが,それはそれで何とか理由をつけるのだろう。結局,両者(生態学者と生理学者)の溝が埋まることはない。
私たちが多くの実験をして得たデータをもとに,生態学関係のジャーナルに出したとしたら,「こんな記載的なデータは要らない,左右性のメリットにかかわる実験をせよ。」というコメントが返ってくるのではないだろうか?左利きであること,右利きであることに特別なメリットがある訳ではないという実験的な根拠を示しても,生態学者を納得させることは不可能である。簡単なことではないかと言うと,研究とは関係のない日常の素行の悪さを取り上げて激しく攻撃してくる。気に入らないことを言われると逆上する者が多い。(人間だけでなく,すべての動物の持つ基本的な性質なのだろう。)周囲の人たちは,そういう者をよく知っていて,うまくなだめすかして場を収める。私は空気全然読めないので,逆上した者の餌食になりやすい。
議論してもかみ合うことがないのであれば,かみ合わない原因を突き止めて対処する方がよかろう。要約すれば,生態学者は生態学的に見て有利とか不利とかを垣間見ることができれば,生理学的なメカニズム(physiological mechanism)は問わないのだろう。だから,生態学者の言う「実験」は,こういうことではないか?「右利きが有利であれば,右手の力が左手の力よりも強いはずである。左利きは右手の力より左手の力が強いはずである。それぞれの手のpinch powerを測定し,統計的検定を行う。左右の力の間に有意な差が認められれば,やはり右利きは左利きに比べて餌をとるのに有利だ。」という論法である。論理的には落ち度がないように見える。
見かけ上であっても,人間は論理的に落ち度がないと分かれば,気持ちが高揚して(舞い上がるとも言う),他人の意見,特に批判的な意見には全く耳を貸さなくなるのだと思う。だから,カニの左利きは,右の鋏脚が取れた結果生じると主張しても,無視されるか,激しく叱責される。多くの生態学者が生理学的メカニズム(physiological mechanism)に抱く嫌悪感は,単なる感情的レベルを越えている。
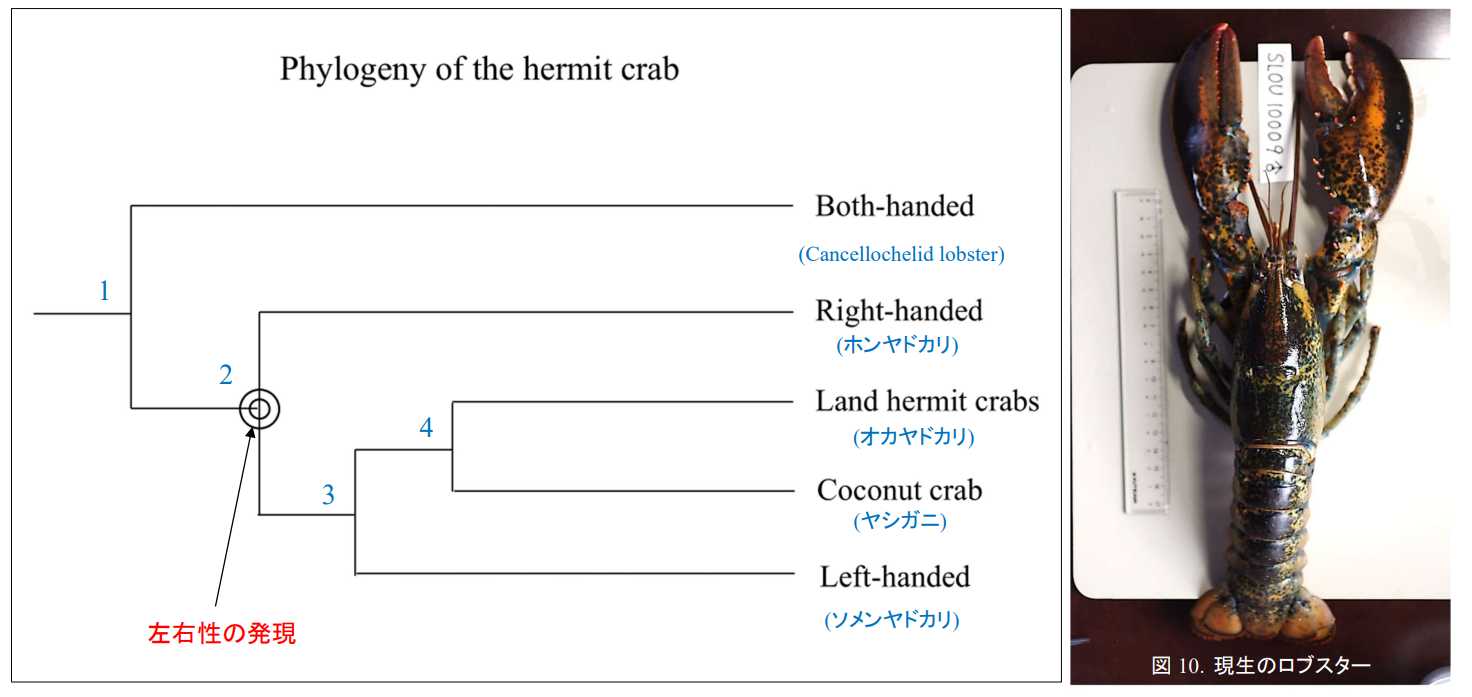
図9.右利きヤドカリと左利きヤドカリの進化。(1:利き手の分化。2:右利きヤドカリと左利きヤドカリの分岐。3:オカヤドカリと左利きヤドカリの分岐。4:オカヤドカリとヤシガニの分岐。ヤドカリはおそらくサンゴ礁原に進出したロブスターの祖先から分かれたと思われる。ヤドカリを生み出したロブスターは,ジュラ紀に絶滅しているだろう。ヤドカリの祖先は貝殻に逃げ込んだので絶滅を免れた。(カニはカムフラージュしたり,逃げ足が速かったりして絶滅は避けられた。)
ガザミの鋏脚(第一歩脚)を構成する各節(掌節,腕節,長節,座節など)を,右と左で大きさ(size)を比較すると,基部の方では左右の節で大きな違いはないが,先端の2節(指節と掌節)で左右に大きな違いが出る。このことから左右性は,液性(内分泌物質)の調節を受けて発現することが予想される。甲殻類の鋏脚(かんきゃく)の外骨格(exoskeleton)を作るメカニズムは,大方わかっていると思うが,左右の節の大きさを調節する機構は,わかっていないと思う。カニ類の胸部下面,厚いクチクラの内側には大きな神経節(胸部神経節)がある。神経節の中にはニューロン(神経細胞)の他に神経分泌細胞(neurosecretory cell)があり,それぞれの細胞から軸索が歩脚の各節に延びている。カニが脱皮するときに,神経節の左右にある神経分泌細胞から歩脚各節の大きさを調節する活性因子が軸索を通して放出され,その因子が鋏脚各節の大きさの調節に関わっていることが予想される。この活性因子の分泌量が左右で異なるのだろう。
また,左右どちらの鋏脚が大きくなるかは系統と密接に関係している。右利きと左利きがいつ分かれたかも明らかにできるだろう。
<調査,写真撮影,原稿執筆の基本情報>
・調査研究者:三枝誠行(生物多様性研究・教育プロジェクト常任理事)。
・写真撮影: 鳩間島の写真は,2024 年 6 月 30 日に三枝誠行が撮影した(Canon EOS 7D, SONYRX10Ⅲ,SONY HDR‒CX680)。
・原稿の執筆:三枝誠行・増成伸文(生物多様性研究・教育プロジェクト)
・編集:生物多様性研究・教育プロジェクト。
・記事の英文題目(Title of the article)。Saigusa M., and N. Masunari (2024) The coconut crab in the subtropical island, Hatoma-jima: cheliped laterality in
the land crabs). In: Research Reports of the Biosphere Project, Section Ⅱ. “Offense and Defense in Decapod Crustaceans”, 2024‒No. 3.
・文献としての表記:三枝誠行・増成伸文(2024)鳩間島のヤシガニ。生物多様性研究・教育プロジェクト Research Reports Ⅱ. 攻撃と防御の十
脚甲殻類 2024‒No. 3。
<参考文献>
- Amati L., R.M., and J.-P. Zonneveld (2004) A new family of Triassic lobsters (Decapoda: Astacidea) from British Columbia and its phylogenetic context. J. Paleontol. 78 (1): 150-168. DOI:10.1666/0022-3360(2004)078<0150:ANFOTL>2.0.CO;2
- Bracken-Grissom, H.D., S.T. Ahyong, R.D. Wilkinson, R.M. Feldmann, C.E. Schweitzer, J.W. Breinholt, M. Bendall, F. Palero, T.-Y. Chan, D.L. Felder, R. Robles, K.-H. Chu, L.-M. Tsang, D. Kim, J.W. Martin, and K.A. Crandall (2014) The emergence of lobsters: phylogenetic relationships, morphological evolution and divergence time comparisons of an ancient group (Decapoda: Achelata, Astacidea, Glypheidea, Polychelida). Syst. Biol. 63: 457–479. https://doi.org/10.1093/sysbio/syu008
- Burggren, W.W., and B.R. McMahon (1988) Biology of the Land Crabs. Cambridge University Press, Cambridge.
- Castro, P., and M.E. Huber (2005) Marine Biology (Fifth Edition). McGraw Hill Higher Education, Boston.
- Horst, M.N., and J.A. Freeman (1993) Crustacean Integument. Morphology and Biochemistry. CRC Press. Boca Raton.
- Karasawa H., C.E. Schweitzer, and R.M. Feldmann (2013) Phylogeny and systematics of extant and extinct lobsters. J. Crust. Biol. 33: 78‒123. DOI: 10.1163/1937240X-00002111
- Masunari, N., M. Hiro-oku, S. Dan, T. Nanri, M. Kondo, M. Goto, Y. Takada, and M. Saigusa (2015) Chela asymmetry in a durophagous crab: predominance of right-handedness and handedness reversal is linked to chela size and closing force. J. Exp. Biol. 218: 3658–3670. https://doi.org/10.1242/jeb.120196
- Masunari, N., K. Sekiné, B.J. Kang, Y. Takada, M. Hatakeyama, and M. Saigusa (2020) Ontogeny of cheliped laterality and mechanisms of reversal of handedness in the durophagous Gazami crab, Portunus trituberculatus. Biol. Bull. 238: 25‒40.
- Richter, S., and G. Scholtz (1994) Morphological evidence for a hermit crab ancestry of lithodids (Crustacea, Decapoda, Anomala, Paguroidea). Zool. Anz. 233: 187‒210.