
2024年8月27日(火)
はじめに
日本列島の泥干潟には,十脚甲殻類(Decapoda)のアナジャコ(Gebiidae)が棲んでいる。どのぐらいの種類のアナジャコが分布しているのか,情報は錯そうしているが,従来の系統学(systematics)や分類学(taxonomy)と分子系統学(molecular phylogenetics)を融合させた実証的系統学(empirical phylogenetics)の視点から整理してみたい。アナジャコ類の分布や分類に関する再構築は,どんな理由で分類の錯綜に至ったのか経緯(人為的かく乱)がわかれば,そう難しくはない。
まずは,アナジャコの祖先(ancestor)について。アナジャコは,サンゴ礁原を生息場所とするアナエビ類(Axiidae)から進化したと思われる。・・・だが,いつどんな種類のアナエビが,サンゴ礁の泥干潟でアナジャコに変わったか,まだわかっていない。
私の現在の仮説を以下に要約する。今から5,000万年近く前(新生代,古第三紀)に始まったインド亜大陸のユーラシア大陸への衝突がきっかけになって,ユーラシア大陸(東南部)の地形(topography)は大きく変化した。結果として,インドシナ半島からオーストラリア大陸の間には,大小さまざまな島が誕生した。
島の周囲にはサンゴ礁(堡礁)が発達し,サンゴ礁原(reef flat)には驚くほど多数の海産無脊椎動物(marine invertebrates)が進出し,サンゴ礁を中心とした熱帯生態系が出現したと考えられる。同時にアナエビ類もサンゴ礁原に進出し,サンゴ塊の隙間に入り込んで生活していたと思われる。
島々は,形成されてからしばらく経つと,土壌の風化が始まる。自然災害(頻発する地震や大雨による洪水)も加わり,陸上からの泥の流出によって,サンゴ礁原の環境が大きく変化したのではないだろうか?サンゴ礁原に堆積する泥は,多くの種類のアナエビを絶滅に追いやったに違いない。一方で,少ないが,新しくできた泥干潟(mud tidal flat)に適応したアナエビ類もいたはずだ。それがアナジャコ科(Gebiidae)に変わったのだろう。
系統学的に「科レベル」の進化となると,1千万年とか2千万年かかるだろう。アナエビ類からアナジャコ(科)への移行は,新生代新第三紀(中新世)に始まったと予想される。小さな突然変異が積み重なり,長い間に今の形態に変わったのだろう。一方,アナジャコになってからは急速に多様化が進んだと思われる。
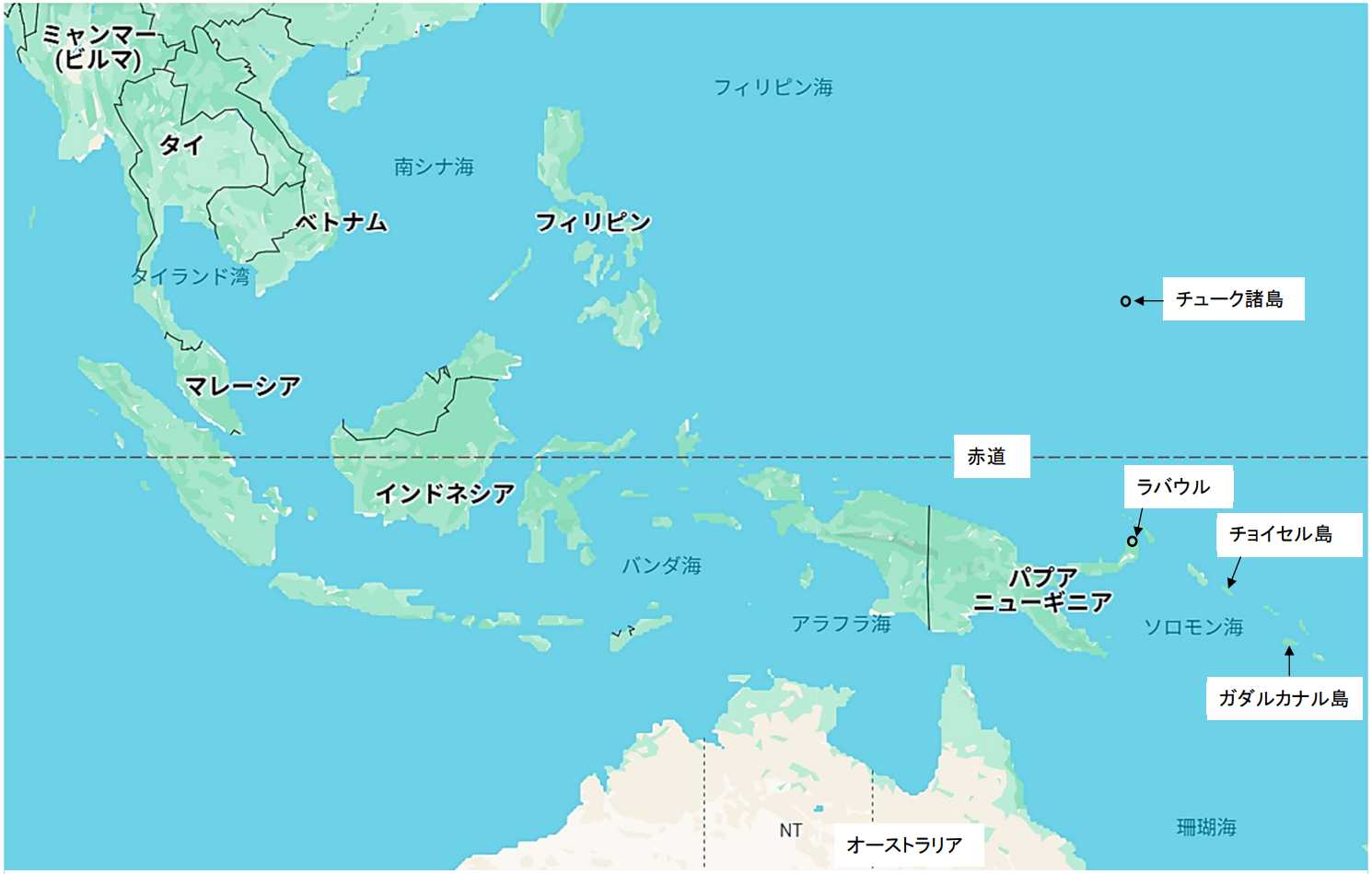
図 1.西太平洋の熱帯域に広がる島々。世界中で広い範囲にこれだけ多くの島がひしめいている海域は珍しい。島々は,海底の堆積層が隆起して海面に現れたのだろう。地層は,泥岩,砂岩,礫岩などで構成されている感じがする。また,南太平洋には環礁(atoll)が多い。環礁ができるということは,島々の地形が大きな速度で変化していることを意味する。一度訪れてみたいが,お金がないのと,多くの島々は治安に問題がありそうな気がする。標本も移送不可能だろう。

図2.ソロモン諸島のチョイセル島(Choiseul Island)東端にある小さな島のサンゴ礁原。サンゴ礁原の深さは,満潮時で2~5m。干潮時になると干潟になる所も多いだろう。アナエビ類は,サンゴ礁原のサンゴ塊(生きているサンゴの塊もあれば,サンゴが死んだ後の塊もある)の隙間に入り込んで生活している。とんでもなくきれいな色をした種類が多いが,採集は難しい。海岸には陸上から泥が流れ込んでいるのがわかるだろう。サンゴ塊の上に泥が堆積すると,サンゴの群体は死滅する。ポリプの口に泥が詰まってしまうためだろう。アナエビも,泥を処理できる種類は少ないと思われる。しかし,一部のアナエビは,顎脚や鋏脚に生えた剛毛の束を使って,水に浮遊する泥を穴の外に捨てられるように進化したと思われる。アナジャコになると,水中に浮遊した泥を顎脚や鋏脚に生えた剛毛の束を使って選り分け,シルトやクレイを食べている。アナエビからアナジャコに至る際の食性変化は,1千万年は十分にかかりそうな気がする。
<質問> 新生代第四紀にサンゴ礁原の泥干潟で誕生したのは,アナジャコ科(Gebiidae)だけか?
サンゴ礁原には,驚くほど多様な生物が生活している。琉球列島の泥干潟を見ると,河口域では確かにアナジャコが優占種になっているが,軟泥層が堆積している干潟ではヒメシオマネキが見られる。底質が砂利になると,とんでもない数のハクセンシオマネキが現れる。泥を掘ると,ユムシ(環形動物),ゴカイ(環形動物),ハサミシャコエビ(十脚甲殻類)を採集できる。海岸の泥干潟では,カニ類を始めとして多くの種類を見ることができる。
マングローブ干潟には,何種類かのシオマネキの他に,キバウミニナ(軟体動物),シレナシジミ(軟体動物),ミナミトビハゼ(脊椎動物)が生息している。さらにヒルギ林の中には,多くのカニ類に加え,背の高い塚を作るオキナワアナジャコがいる。
河口域の泥干潟やマングローブのヒルギ林に生息する動物は,独特の形態を持つ種類が多い。サンゴ礁原では,浅潮下帯(shallow subtidal zone)に住む動物と同じか,非常によく似た形態の動物が多い。陸上から流入する「泥」は,有機物を多く含んでおり,生物の十分な栄養源になる。一方,泥(シルトやクレイ)の粒度は小さく,水中に浮遊する密度が高くなると,生物の生存に大きな支障をきたすだろう。例えば,サンゴ礁に泥が流れ込むと広範囲のサンゴが死滅する。だから,サンゴ礁原に生息する生物にとって,泥は環境汚染物質と言える。
新生代第三紀のあたりから,河口域だけでなくサンゴ礁原にも大量の泥が堆積することが多くなり,最初のころは,泥の堆積によってサンゴ礁原の生物相は大きく破壊されただろうが,泥が堆積する環境でも生きられる生物が出てきたのではないだろうか?泥干潟への適応は,見かけほど簡単ではなく,1千万年はかかったかもしれない。その間に形態(morphology)の変化が進んだ生物がいたことは十分に予想される。カニ類ではシオマネキが良い例になる気がする。
カニ類の場合には,もともと砂利とか砂の上を歩く生活をしており,ガザミのように軟泥層の表面でも昔と同じ生活ができた種類がいるだろう。シオマネキやミナミコメツキガニは,祖先がどんなカニだったか不明だが,泥干潟への進出にともない,形態や行動も大きく変化したのだろう。頭胸部や鋏脚の構造からみて,アナジャコはサンゴ礁原に住むアナエビから進化したと考えて間違いないと思う。泥干潟という新しい環境に適応する過程で形態も大きく変化したと考えられる。
アナジャコの場合には,泥の中に巣穴を作るという性質がある。捕食者は巣穴の入り口で追い払うことができる。巣穴の中で体を迅速に移動させられる。泥の粒子はエサにできるが,砂粒になると顎脚に生えた剛毛の束を使って排除する性質を獲得している。泥の中で生活するのに役立つ形態変異をもたらす突然変異遺伝子が蓄積されたと思われる。泥干潟という新しく広がった環境に適応するためには,アナエビ(Axiidae)という「形態」のままでは,無理があったということかも知れない。(突然変異はしばしば形態の変化をともなう。)しかし,アナエビからとんでもなく形(form)を変えてしまうことも難しかったのだろう。
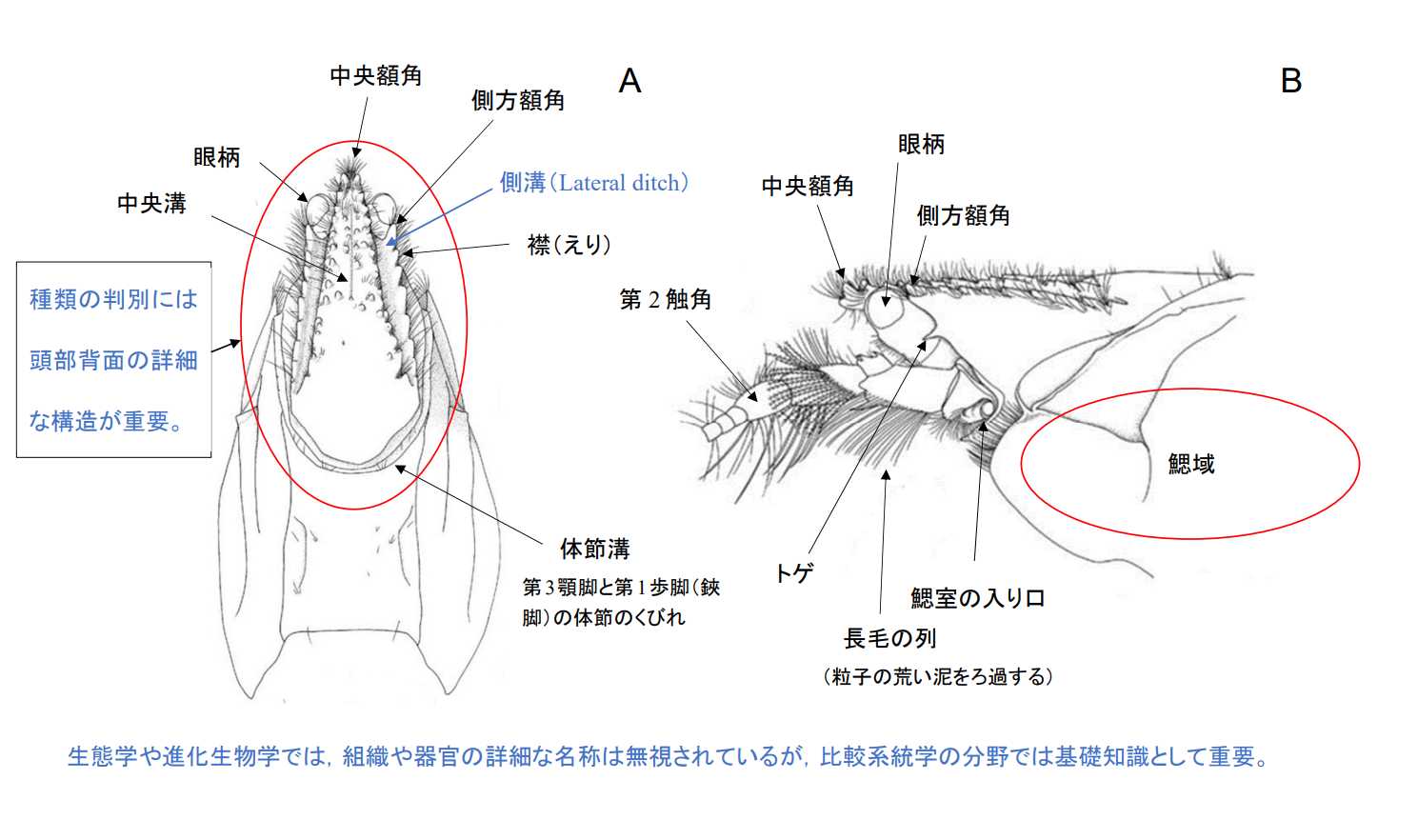
図3.アナジャコ頭胸部の形態。A: 頭胸部背面,B: 頭胸部側面。十脚甲殻類は,攻撃と防御に頭部の器官(角やトゲ)と強力な鋏脚(第1歩脚)を使用する。そのため頭胸部の背面と鋏脚の外骨格(exoskeleton)は非常に厚く,また硬い構造になっている。頭胸部背面や鋏脚には,種固有の構造が発現していることが多く,重要な分類基準となる。ただし,分類の基準は分類学と比較系統学ではいつも同じという訳ではない。(Hirano and Saigusa 2008, Figure 2A and 2Bを転写)
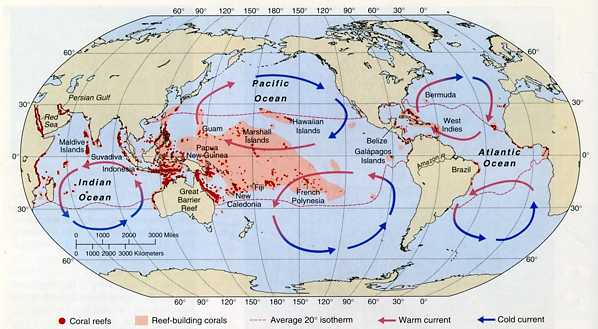
図4.サンゴ礁の分布と海流の循環。赤い斑点は,礁(reef)を作らないサンゴ塊を示し,ピンク色で塗りつぶした部分は礁を形成するサンゴ(ミドリイシの仲間)の分布(つまりサンゴ礁)の分布を表している。地球の海洋に分布するサンゴの種類は,ピンク色の海域が最も多く,そこから離れるにしたがって同心円状に(と言っても,横方向にゆがんだ同心円になるが・・・)減少する。インド洋や紅海はサンゴの種類は多いが,サンゴ礁は発達していない。(Castro and Huber 2005, Figure 14.10を転写)
図4を見ると,サンゴ礁の中心は,経度ではボルネオ島からフランス領ポリネシアの間,緯度で言うとニューカレドニアからフィリッピン,グアム,マーシャル諸島にかけての熱帯域にあると言える。サンゴ礁は大陸沿岸には形成されず,熱帯の島々の周囲に見られる。サンゴ礁の形態については,ピンク色の西側半分は堡礁(barrier reef)が多く,東側半分になると環礁(atoll)が多くなる。日本列島のサンゴ礁は,すべて堡礁に分類される。サンゴ礁のパターンとして,裾礁(きょしょう)が区別されている。(「裾」は「すそ」の意味。)裾礁は,概念としてはありと思うが,現実的にこれが裾礁だと言えるサンゴ礁があるかは疑わしい。海洋生物学の教科書でもあいまいな図が置かれている。
日本列島では,暖流(黒潮)の影響で奄美大島以南の琉球列島ではサンゴ礁(堡礁)が見られる。琉球弧は,ユーラシア大陸プレートの下にフィリッピン海プレートが潜り込む反動で,大陸棚の縁が盛り上がり,帯状に隆起してできたのだろう。台湾から奄美大島までの弧状に連なった島々の周囲に形成されたサンゴ礁を,Ryukyu Great Barrier Reefと呼んでも間違いではない気がする。
<生命の起源と細胞の進化>
地球上で最初の生命(シアノバクテリア)が誕生したのが,35億年前。それから20億年近く,原核細胞(光合成細菌や細菌類)の時代が続いた。原核細胞の生存に使われる栄養源(エネルギー)は,自身が作り出すか,他の細胞を捕食・消化によって得られる有機物(organic matter)だったろう。無機物は分解しても大きなエネルギーは得られないのではないか。逆に分解するのにエネルギーが必要という物質が多いのではなかろうか?
エネルギーの本質はよくわからないが,エネルギーあれば,物質は今で存在しなかった新しい構造を作ることができる。構造(structure)は,生物の場合には形態(morphology)と言い換えられる。さらにエネルギーが供給されれば,その構造は破壊(分解)に傾くこともあるだろうし,さらに新しい構造の構築につながることもあるだろう。始原地球では,地表に適当な温度(熱)と水の存在によって,他の星では起こりにくい物質の変化(化学合成と新しい構造の発現)が可能になったのだろう。
結果として,生命(シアノバクテリア)は,自身ができるだけ長く生存し,できる限り多くの子孫(単細胞生物の場合には「同胞」)を増やすという2つの宿命(生命の維持・増殖の原則)を背負うことになったのだろう。また,生命の維持と増殖の原則は,解消するか発展的な方向に進むか,どちらかに分かれるため,自己増殖をする段階(生命)になってから,生命はずっと同じ状態にあったのではない。できる限りエネルギー効率のよい摂食法を巡って,原核細胞同士の間に競争が起きたことは容易に推察される。食べたり食べられたり,くっついたり離れたり,だましたり騙されたり,多様な生き方が可能だったと思われる(原核細胞における三国志時代)。その中で勢力を持ち始めた一派(いくつかの派閥がある)が,真核細胞に発展したとは考えられないだろうか?不思議なことは,原核細胞では互いに潜り込んで生活する進化(共生)は起きたが,多細胞になろうとした系統が見当たらないことである。
競争の原則は,真核細胞になってからも生物界に適用された。わが一族は,分裂後の細胞同士が常に対立関係にあり,決して仲良くなることはないという原生生物もあっただろう。いやいや,我ら兄弟・姉妹は常に仲良く,国難には協働して対応するぞという一派(系統)もあっただろう。原核生物(モネラ界)と同様に原生生物(真核生物として一番最初に地球上に出現)にも多様な生き方あったと思われる(真核細胞における三国志時代)。結果として,細部分裂で増殖した真核細胞同士は接着し,多細胞生物が誕生した。動物界(Animal Kingdom)では,ヒドラ,クラゲ,イソギンチャク,サンゴが含まれる刺胞動物が最初に出現した。
なお,今西錦司氏は新しい生物群について,「生まれるべくして生まれた」みたいな考え方をしていたかもしれない。今西進化説だと,生物の進化が心霊現象の一種にされてしまう。私は生命と生物の進化を「物理・化学の法則」とダーウィンの提唱した「生存競争」という概念を軸に考えていった方が良いと思う。最近現れた「遺伝子の適応度」という概念やヘッケルの生物発生の原則も使える。一番まずいのは,法則の結果として発現した生態学的現象(例:すみわけ)を,生物原則(原因)と取り違えることである。ここがあいまいになっている研究者が多い。
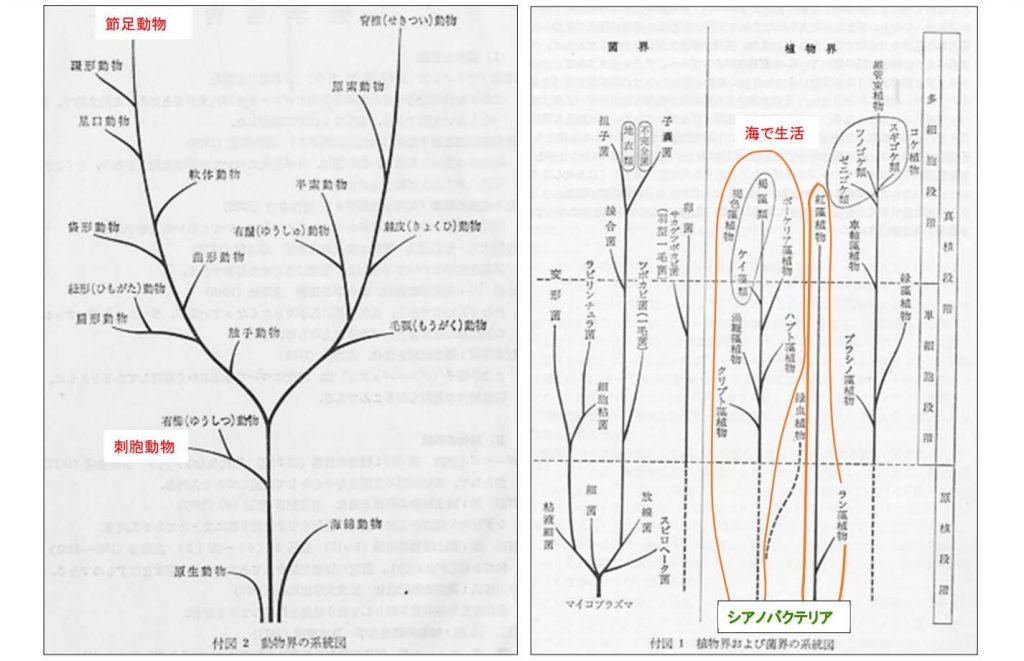
図 5.生物界の系統図。山田・西田・丸山「進化系統学」の付図を転載。生物の進化は,実際にはこんなに一直線状ではない。新しいタイプの生物群(taxon )が現れると多様化が起きる。その中で生き残った群が,次の時代の新しい分類群に発展する。では,古いものはすべて絶滅するかというと,決してそんなことはない。新しいタクソンとは顔を合わせないハビタットに逃避するか,新しい群に寄生して種としての延命を図る種類も多い。
<琉球列島におけるアナジャコの種類と分布>
私は日本列島の干潟に分布するアナジャコの種類は多くないと思う。チョウでは台風に乗ってやってきて,一時的に繁殖する種類(迷蝶)がいる。野鳥では,繁殖地や越冬地から遠く離れた場所に来る個体(迷鳥)がいる。
アナジャコでも,たまたま幼生(larva)が流れ着いて,その場所の干潟で生活していた個体が採集されたという例はあるだろう。分類学的には,そういう個体は大事なのかもしれない。しかし,実際に個体群が分布しているかという話になると,実際に生息場所に行って調べてみる必要がある。
琉球列島に関しても,どの島にどんな種が分布しているのか,よくわかっていないのが現状だろう。
琉球列島に分布するアナジャコ類の分布について,一番正確に記述されたと思われる文献は,Itani(2004)と思われる。伊谷行氏は実際に琉球列島に行き,採集した標本をもとに特定した種類をまとめたのだろう(図5)。ただし,それぞれの種類の図や写真(証拠)がなく,採集された個体の形態の比較ができないため,種の特定ができない。進化系統学では,それは困る。

図 6.琉球列島に分布するアナジャコ(Itani, 2004 から転写)。琉球列島には,Upogebia yokoyai, U. carinicauda, U. pugnax の 3 種が分布しているとされたが,Itani(2004)(博士号の学位審査の際に提出する参考論文?)には,図(スケッチ)も写真もない。宮古島では,与那覇湾と大浦湾で U. pugnax が採集されているが,私はどこの島でも「・・・湾」での採集成績は極端に悪い。また,U. pugnax というのはどんなアナジャコなのかも,イメージがわかなかった。証拠を提示せずに意見を主張しあうと,議論の落としどころがない。
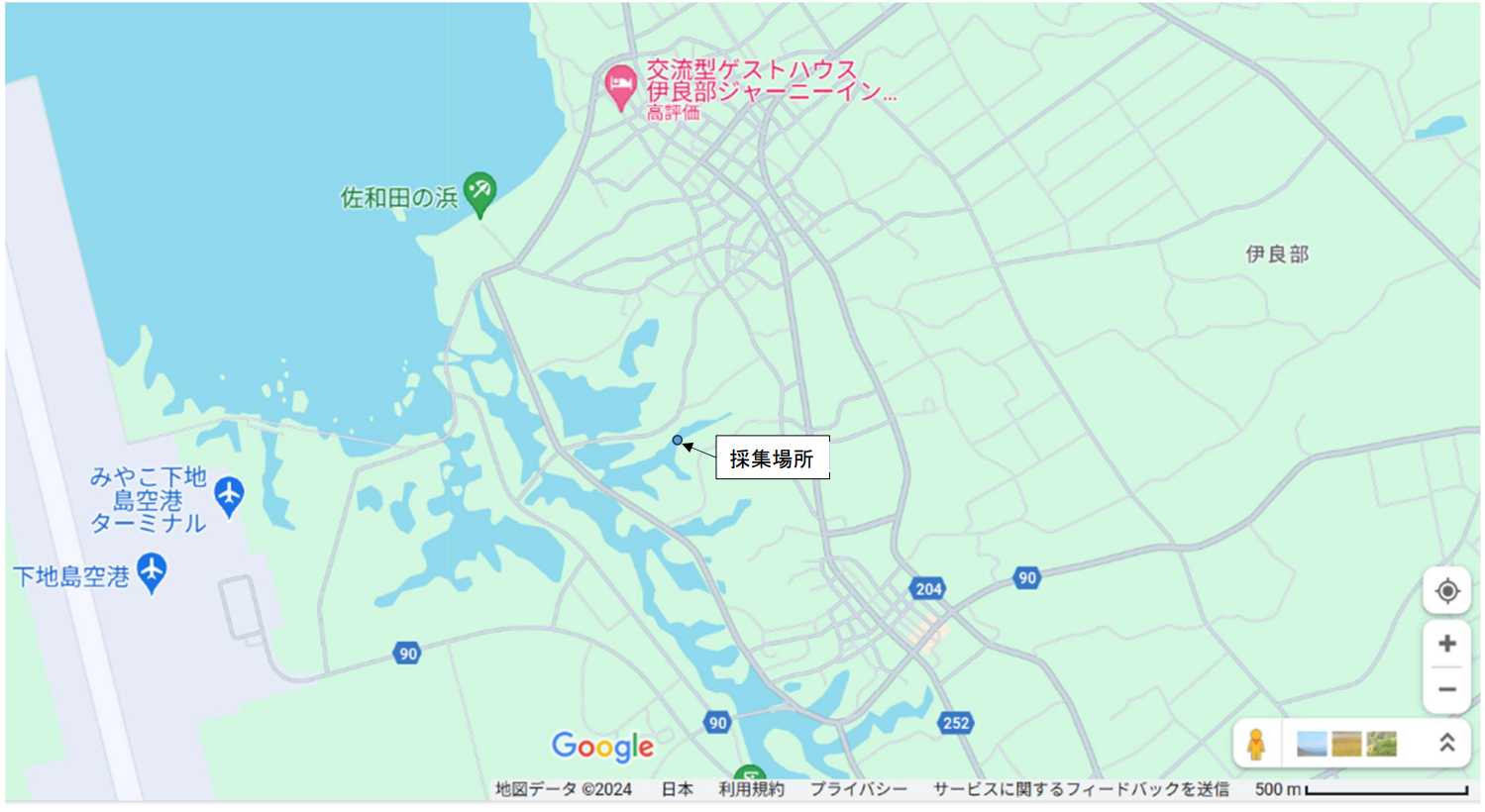
図 7.伊良部島におけるアナジャコの採集場所。伊良部島と下地島の間は,細長い入り江(inlet)となっている。干潮に海岸を含めて入り江に下りてアナジャコの巣穴を探した。巣穴は各所に見つかるが,砂泥底をスコップで掘っても,アナジャコは全然採集できなかった。ただ 1 か所,マングローブ干潟でアナジャコとハサミシャコエビが捕れた。伊良部島と下地島の間の入り江は,双方の島にかかる橋の上から見ると風光明媚だが,入り江に釣りに来る人たちが,車の駐車スペースや干潟にゴミを捨てて行く。全部ではないだろうが,釣りをする人たちは,目的の達成ばかり考えていて,自分たちの行動が環境にどれだけ大きな負荷をかけているかなど考えもしないのだろう。今まで静かだった観光地に,ある時を境に大量に押し寄せ,一日中騒ぎまくり,路上にゴミを放置して帰って行く人たちは多い。バーベキューをして,飲んで騒いで大量のごみを海岸に残す「やから」も少なくはないだろう。ゴミの放置や河川や海岸への投棄は,日本だけでなく,世界中で起きている深刻な環境問題である。

図 8.伊良部島のマングローブ干潟。干潮時になると,入り江に泥干潟が現れる。堆積した軟泥層の表面には,直径 5~6mm の穴がたくさん開いていて,スコップで泥を掘るとアナジャコ,スナモグリ,ハサミシャコエビを採集することができる。軟泥層は,陸上から流出した赤土だろう。

図 9.泥を掘って採集されたアナジャコ(メス)。この時点では種名はわからない。軟泥層は厚く,深さ 30cm 近くあっただろう。長靴が泥に埋まって足を取られるので採集は結構難儀である。しかし,ここの干潟は軟泥層の下は砂地なので,採集はしやすかった。軟泥層の下にサンゴ礫があると全然捕れない。

図 10.伊良部島のマングローブ干潟で採集されたアナジャコ。左がオスで右がメス。オス(左)の鋏脚の外骨格には赤い斑紋(色素胞)や筋がある。オスの胸部と腹部の一部に見られる黄色い部分は中腸腺(栄養分を貯蔵する器官で,消化液は分泌しない)。腹部の黒い帯は腸管。メスの胸部から腹部全域に見られる緑色の器官は卵巣である。宮古島のアナジャコの受精卵はきれいな緑色をしている。アナジャコのメスでは,内臓容積の 8 割から 9 割が卵で占められていると思う。一年に 2回は産卵すると思われる。体の全容積の 6 割か 7 割を生殖腺が占めていると思われるが,どうしてこんなにいびつな生命体になってしまったのだろうか?そんな不自然な体になってまで子孫を増やそうとすることの「大義」(cause)はどこにあるのだろうか?
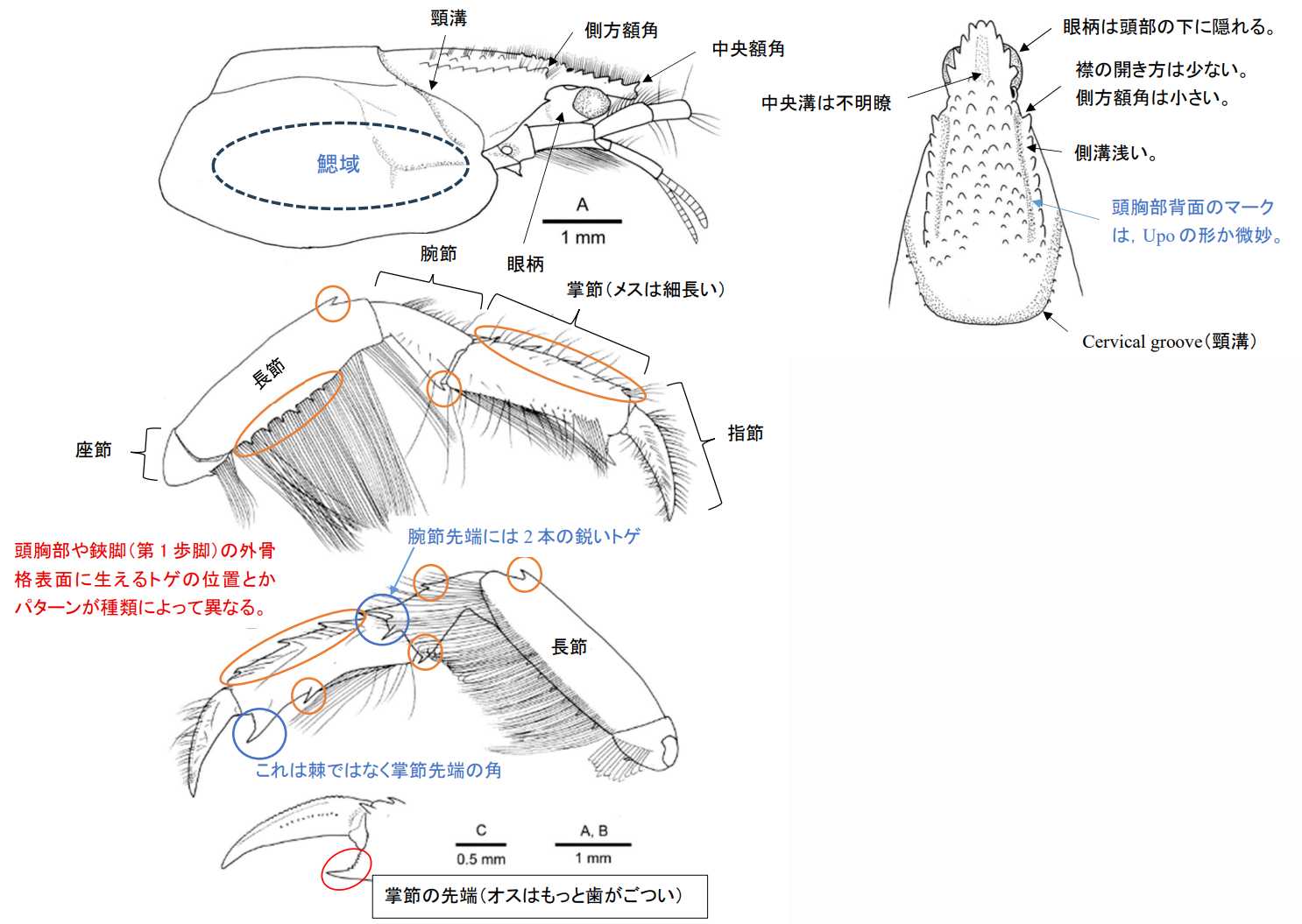
図 11.Upogebia pugnax の外部形態。
Komai (2020) Figs. 1 and 2 を転写。スケッチしたのは西表産の個体。オスと書かれているが,メスである。
西表島ではコブシアナジャコは,湾内の潮間帯から浅潮下帯で採集されるようだ。鹿川湾でダイビング中に採集したというのは驚きである。まさかそんなところにいるとは・・・。
仲間川の河口(大富と大原)にも,大潮の干潮時には干潟が出現する。本種はそんなところで採集されるようだ。私も何度か仲間川の河口(干潟)で採集したことはあったが,Upogebia pugnax は一度も捕れたことがない。私は海岸での採集は極めて不得手である。
Pugnax は,宮古島でも海岸で採集されているようだが,私は海岸で捕ったことがない。西表島では,河口域(マングローブ干潟)には,Upogebia yokoyai (正確には Clade C と D)が分布し,U. pugnax はいない。Itani (2004)も,おそらく私が採集したのとよく似た環境(干潟)で採集していると思われる。西表島では pugnax はサンゴ礁原の泥干潟にいるようだ。
一方,宮古島のマングローブ干潟では,pugnax ばかりが採集され,yokoyai の気配は全くなかった。
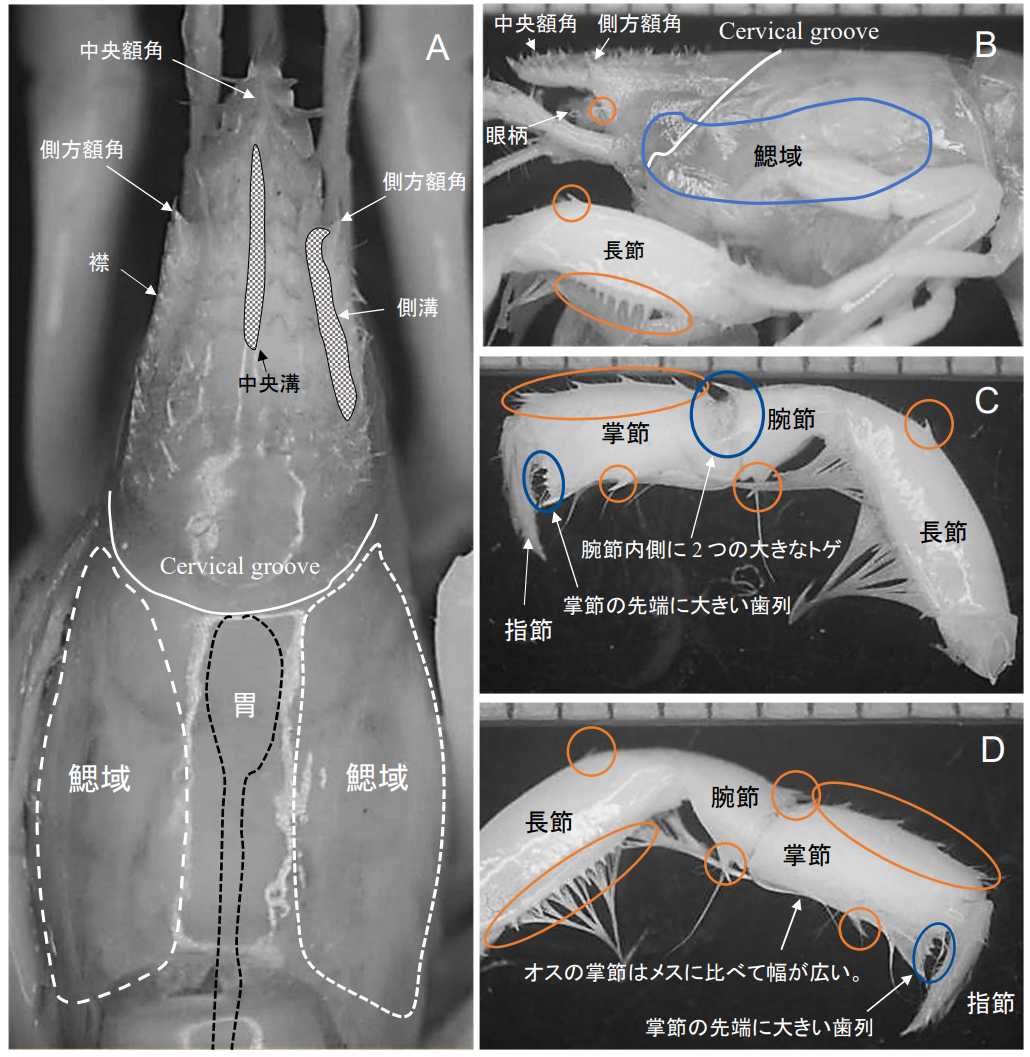
図 12.Upogebia pugnax (オス)。伊良部島産。
A: 頭部背面。 B: 頭胸部側面。
C: 右鋏脚(第 1 歩脚)内側。
D: 右鋏脚(第 1 歩脚)外側。
十脚甲殻類は,各体節に一対の付属肢(pleopod)が並んでいる。胸部(thorax)の付属肢は歩脚(pereiopod),腹部(abdomen)には腹肢(遊泳肢)が並んでいる。
歩脚に関しては,雌雄で数(5 対)と形態に違いはないが,腹肢は雌雄で大きな違いがある。メスでは,第 1 腹肢から第 5 腹肢が見られる。第 2 腹肢から第 5 腹肢には,長毛(long setae)
が生えているが,第 1 腹肢は細く短い棒状になっている。
オスの場合には,第一腹肢は欠損しているが,時々第 1 腹肢の生えた個体も混じる。第 2腹肢から第 5 腹肢の形状はメスと同じ。
生殖口に関しては,オスは第 5 歩脚の付け根に雄性生殖口(精包の出口)があり,メスでは第 3 歩脚の付け根付近に雌性生殖口(受精卵の出口)がある。
しかし,オスにもかかわらず雌雄両方の生殖口を有する個体がいる。(U. major ではオスにも卵巣が見られる。)メスにも雄性生殖口が見られる個体がいる。(精巣はないと思う。)
アナジャコの場合には,種によっては雌雄の形態が大きく異なる。雌雄をよく調べないまま分類すると,とんでもない間違いを犯すだろう。
図 12 の個体はオス。Itani (2004)が発見し, Komai (2020)が記載した Upogebia pugnax に間違いないだろう。しかし,Komai (2020)が記載した個体とはいくつか異なる特徴がある。
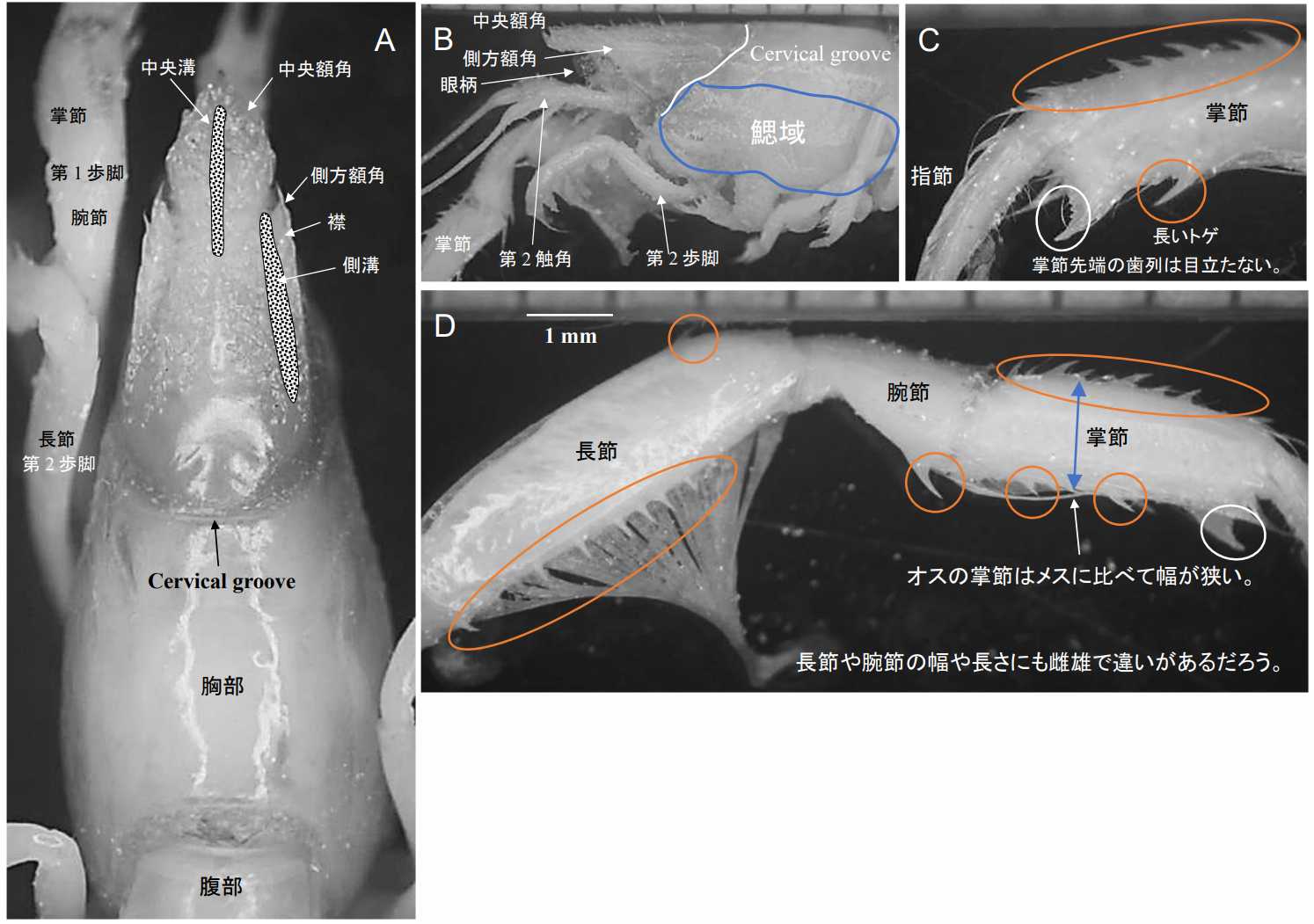
図 13.Upogebia pugnax(メス)。A: 頭部背面。 B: 頭胸部側面。C: 右鋏脚(第 1 歩脚)内側。D: 右鋏脚(第 1歩脚)外側。
Upogebia pugnax は,オスとメスで色彩も違うが,第 1 歩脚の形態も大きく異なる。Komai (2020)が描いたの(図 11)は,図 12 に示した個体(オス)ではなく,図 13 に示した個体(メス)だろう。アナジャコの付属肢(appendage)雌雄の違いに関しては,いずれ SEM(走査型電子顕微鏡)の写真を添えて別な機会に紹介したい。ここで指摘したいのは,雌雄の違いが大きい種では,メスをオスと取り違えて記載すると,本当のオスが発見されたときに新種(亡霊種)として認定せざるを得なくなる。分類に混乱を引き起こすので注意。
<ヒトの進化からアナジャコの進化を考える。>
【人の進化について考える基礎となる記述的データ】
高等学校の教科書「世界史」に記述されたヒトの進化は,人類学的視点から次のように要約される(三成 2022)。
化石の証拠によって,人類(human kind)が誕生したのは今から500万年前の新生代・新第三紀の鮮新世と考えられる。誕生した場所はアフリカ大陸のサバンナか?
最初の人類は,人類史の中では猿人(ape-man)に分類される。動物学の視点では,猿人の学名はAustralopithecines(科?)で,以下の5つの属(genus)で構成されている。Australopithecus, Paranthropus, Kenyanthropus, Ardipithecus, およびPraeanthropus。なお,それぞれの属はどのぐらいの数の種(species)で構成されているかは不明。1属1種の可能性もある。
次は原人(early man)で,180万年前に出現した。Homo antecessor, Homo erectus(ジャワ原人,北京原人を含む), Homo ergaster, Homo heiderbergensis(ハイデルベルグ人),Homo floresiensisなどが含まれる。原人の段階で,地球上の広い地域に分布し始めていることがわかる。
さらに約20万年前に出現したのが旧人(Archaic humans)で,Homo neanderthalensis(30万年前~4万年前),Homo rhodesiensis(30万年前~12.5万年前),Homo heiderbergensis(60万年前~20万年前)が含まれる。Homo antecessor(120万年前~80万年前)を含める場合もあるようだ。種名が原人とダブっている種類がいる。原人と旧人は,形態で区分しにくいのだろう。新しい種(species)が誕生するには,300万年はかかるという証拠がある(Futuyma 1998)。もしその証拠が間違っていなければ,30万年から120万年という期間は,新しい種の誕生には短すぎる。
最後は約4万年前に出現した新人(Homo sapiens)である。新人には現代人の他に,クロマニョン人が含まれる。ヒトは,猿人・原人・旧人・新人の順に進化してきた。文字が生まれたのは,紀元前3千年ごろ。文字をもつ以前は先史時代(紀元前3千年以前),文字を持ってからは歴史時代(紀元前3千年から現代までのおよそ5千年)と呼んでいる。先史時代についてはまだ謎が多い。しかし,考古学,人類学,民族学の発展によって,ヒトの系統(phylogeny)について,真実の姿が明らかになりつつある。
私(三枝)の個人的意見として,ヒトの進化を説明するときにひとつ大事な注意点を述べておきたい。人類学的ヒトは,猿人・原人・旧人・新人に分類される。それぞれの分類群(taxon)の形態を比較すると,原始的な特徴は猿人が一番多く,新人が一番少ない。だからヒトは猿人・原人・旧人・新人という順で進化したと考える人は多いだろう。道徳観念の強い人,実証的なデータの取得よりお話を作ることが好きな人に好まれる仮説である。
一方,原人と旧人の区分はあいまいなところがある。新人は旧人から進化したのではなく,本当は原人の一部から(旧人とは別のルートで)進化した可能性は十分にある。生物進化では,むしろそんな方の経路(系統)が普通である。

図 14.Upogebia pugnax の脱皮殻。
アナジャコは,泥干潟に作られた巣穴に四六時中こもっていると思われているが,時々外に出ているのではないだろうか?大潮のころ,潮が上げてきたときに泥干潟の上を歩き回ると,たくさんのアナジャコが泳いでいることがある。
アナジャコが生息している泥干潟では,泥の上に脱皮殻がたくさん落ちている。どの脱皮殻も壊れている感じがしない。アナジャコは,脱皮する際には巣穴の入り口を広げて泥の上に出て,泥の上で脱皮してからまた元の巣穴に戻り,巣穴の入り口を元通りに狭めるのではないだろうか?
巣穴の中で脱皮した場合には,外に押し出すときに壊れてしまうように思われる。また,巣穴は一匹通るのがやっとの細い径なので,巣穴の中で脱皮するのは難しいような気もする。生殖期になると,オス・メスともに巣穴を出て,オスからメスに精包の受け渡しが行われるのだろう。私はまだ観察したことはない。
【なぜ人類史における「猿人・原人・旧人・新人」の区分は使われないのか?】
三成(2022)によれば,今日では「猿人・原人・旧人・新人」という名称は外国ではほとんど使われない。特に「旧人」という名称は全く使われていないとのこと。理由は,ホモ・サピエンス(新人)が,ネアンデルタール人(旧人)より古い時代に棲息していたことがわかったからである(溝口2011:p.39)とある。
生物の進化は,進化段階の一般的な定義と系統学上の実際の位置(status)という2つの側面で説明される。ヒトの進化をわかりやすく説明するのであれば,猿人・原人・旧人・新人という進化段階を仮定することができる。進化の「お話」として,猿人・原人・旧人を経て新人に至ったとする考え方(生態学的説明)は,ある意味正しいのであろう。しかし,比較系統学的に調べてみると,生態学的な説明はインチキだったということは普通にある。
人(human beings)は,ファンタジー(妄想)に容易に騙されやすいという欠点がある。生態学的な説明(分類学も同じ)は,検証が必要な仮説であるにもかかわらず,いつの間にか新しい知見や事実として人々の心の中に定着させてしまうという(宗教的)魔力を秘めている。おとぎ話として完結していれば,検証する必要はないし,検証してもらっては困るのである。
おとぎ話を検証するためには,生態学的説明と生態学者の言う記載的なデータ(詳細な観察,分子系統図,化石の記録)を比較してみればよい。大概の生態学的説明の矛盾が明らかになる。
問題は,生態学的説明に証拠を示して「問題あり」となったときに,果たして生態学者(分類学者も同様)は,自分たちの間違いを素直に認めるかということである。例えば化石の証拠を示して「ホモ・サピエンス(新人)が,ネアンデルタール人(旧人)より古い時代にいた。」と公に主張すれば,水増し仮説にご執心の方々(昔の大学紛争のリーダーのような人たち)の手にかかると,こんな説明はすぐに論破されると思う。
ラマルクの場合にも同じことが言える。「生物は進化する」という感覚は,ラマルクからすればひとつの仮説にすぎないことだった。何らかの手段で検証して,仮説は確からしいという結論になればそれでよいし,ちょっとありえへんという結果になれば,それでも納得したであろう。しかし,狂信的実証主義者により頭ごなしに否定され,公私にわたって執拗な嫌がらせを受け続けた。
偏狭な実証主義者の考え方,すなわち新しい技術を使ってできる限り正確な記述的データをとることが生物科学の発展につながるという主張は,わからない訳ではない。しかし,詳細な記述的データのみを羅列したら,聞いている方(特に生態学者や分類学者)は退屈で仕方ないと思うだろう。真実が目の前にあるのに,くどくどとつまらない記述的データを出して時間を浪費するなと,お怒りの言葉を発するだろう。国会の答弁で,そばにいる官僚が記述的なデータ(証拠)を説明しだすと,激怒する政治家(特定の人が頭に浮かぶ訳ではない)と似ている。
一番の問題はどこにあるのだろうか?人は誰しも,ファンタジー(ポジティブ,ネガティブ両方ある)に容易に騙されるという性質を持っていることだと思う。もちろん個人差はある。何かのきっかけ(刺激)で脳が興奮すると,妄想が生まれ,今までと違った行動が発現しやすくなるのかもしれない。
人の脳はストレスにも弱い。具体的な例を挙げて申し訳ないが,自分の思ったように周囲が行動できないことに腹を立て,あれこれ当たり散らす方(例えば,豊田真由子さま)は,私自身も含めてしばしば目にする。小保方晴子さんも,研究業績という重圧の中でネガティブな妄想が生まれたのではないか?
生物はみな「後悔先に立たず」という時間原理の中で生きている。時々立ち止まって後ろを振り返る余裕がないと,一生を棒に振ることになりかねない。その意味では,時々授業をさぼって旅行に出かけたり,虫取りに行くなんていいかもしれない。
自然科学の世界であれば,仮説と事実を混同しないこと,および客観的な方法によって仮説は検証されるという認識を共有することは大事かと思う。どの時代にも図上演習が好きな人たちがいる。そういう人たちは検証と称して,偏向した思考(思想)によってシミュレーションを捻じ曲げる傾向が強いので警戒が必要。
また,有効な検証方法の開発(化石のさらなる発掘,分子系統学的手法の改良,比較系統学的解析の導入,統計的検定の導入など)は,生態学,分類学,進化生物学の発展(思弁的生物学から実証的生物学への転換)に大きく寄与するだろう。
(参考文献)
三成美保(2022)先史時代の世界(人類の起源・移動地図・年表)。(https://ch-gender.jp/wp/?page_id=5378)

図 15.伊良部大橋。奥に見えるのが伊良部島。橋の中央に focus が合っているため(ズームレンズを使用),橋の傾斜が増幅されて,すごい急斜面を上り下りする感じになる。実際には斜面の傾斜は写真の半分ほどだろう。伊良部島は,今から 200 万年前にサンゴ礁原が隆起してできたようだ。
系統学的視点での属と種の再編成(案)
・ヒトの進化をアナジャコに適用して,比較系統学的視点から今まで報告されたアナジャコの属や種を再編することが目的。私は分類学者と違って,新種を記載することには興味がない。生物進化を比較系統学的視点で明らかにしたい。
・今まで報告されたアナジャコの属や種に関する文献を調べつつあるが,属や種の認定の基準(standard)が不明な論文が多いために,手持ちの個体との間の比較(comparison)ができない。(特にスケッチ同士の比較は難しい。)もちろん,一定の基準(学会で認定されているとかの基準ではなく,個人の考えている基準)に基づいて属や種の同定に関する仮説をパブリッシュすることは大いに結構なことだが,比較ができないと私の方は研究を進められない。そもそも誕生してから100万年もたたないような短い時間で,あれだけ形態の多様化が進んだと言い切る証拠はどこにあるのだろうか?手持ちの標本と比較しながら,アナジャコの属や種の再編が必要である。
・Upogebia属の和名は,ヒメアナジャコ属とする。(「ひょうたんゲビア属」という日本語は印象が良くない。)従来ヨコヤアナジャコとされていた種(Upogebia yokoyai)は,生息域の異なる2つの種に分割できる。Clade A, E, Fは Upogebia continentalis(キタヒメアナジャコ)Clade B, C, DはUpogebia kariyusi(ミナミヒメアナジャコ)にまとめられる。U. yokoyaiという名称は消滅。
・Upogebia continentalis(分類学ではUpogebia yokoyai)のクレードFは,本州沿岸に局地的に分布している。既に報告されている種(Komai 1999)と同じ可能性がある。
・宮古島で採集されたアナジャコ(図11と12)は,系統からみると一番古そうである。分布は,図4に示したサンゴ礁原の分布とほぼ一致しているのではなかろうか?・・・ということで,属をUpogebiaからArchaeogebia(ムカシアナジャコ属)に変更。Upogebia pugnaxはArchaeogebia pugnaxとなる。和名は,私はアカモンムカシアナジャコと呼びたい。Upogebia sakaiiはArchaeogebia pugnaxと同じなので統合。
・Upogebia miyakei(Hirano and Saigusa 2008)(図3)は検討中。Upogebiaの一番古いタイプなのだろうか?いずれにせよ miyakeiという種名は変更される。人名がつくと,比較系統学という学問から離れて,百人一首のカルタ遊びという印象が強くなり,比較系統学の分野では使えない。それと種名の中に,対象とする種の情報を入れておきたい。Upogebia miyakei の場合にはUpogebia antecessorisはどうか?Antecessorisは「元祖の」という意味。
・Upogebia carinicauda(Itani 2004)は,私は今まで採集していない。Upogebia rupicolaに酷似しているという報告がある(Komai 2005)。頭部の構造(マーク)から,Upogebiaの系列ではないように思われる。検討中。
・砂岩や泥岩に住むイシアナジャコ類(Stone gebiids)は,Stonogebiaという属にまとめるのは無理。(3つぐらいの系統が入り込んでいる。)再編成について現在検討中だが,Upogebia saigusaiについては属名・種名ともに変更する。本種をStonogebia属かSpinogebia属とする可能性もある。Iriomotensis, spinidactylus, semicircula (Saigusa et al. 2014)はUpogebia属にまとめられるか検討中。
・Upogebia neogeniiは,平野(personal comm.)によれば,形態的に区別できるとのことだが,私にその能力があるかどうか?形態的に区別できたとしても,Iriomotensis, spinidactylusから生殖隔離が起きていない可能性もある。その場合に別種と認定するかは,定義の問題になる。分類学者は,生物の種を分けることに命を懸けている。分けないと気が済まない性格の持ち主が多いようだ。
・東南アジアの島々の干潟に住むアナジャコ類に関しては,今よりも種類数は大幅に減少することが予想されるが,博物館等に所蔵されている標本を調べてみようとは思わない。私の方は,分類学者の中では常識とされる主張,例えば大陸沿岸の種との比較が常識であるとする主張(習慣),はよく理解できない。
・生息場所の環境についての情報が必要だが,採集自体が不可能である。ソロモン諸島だけでも難しいだろう。東南アジアの島々を始めとして世界中の干潟にどのように広がったかについては,今の私の技量では答えようがない。
1. 撮影と執筆の基本情報
<撮影> 三枝誠行 <アナジャコの撮影場所> 伊良部島
<撮影機材> 野外ではEOS 7Dと RICOH WG-50。標本の撮影:顕微鏡撮影装置
<執筆> 三枝誠行・増成伸文(NPO法人 生物多様性研究・教育プロジェクト理事)
2.参考文献
- Castro, P., and M.E. Huber (2005) Marine Biology. Fifth Edition. McGraw Hill, Higher Education. Boston.
- Hirano, Y., and M. Saigusa (2008) Description of the male of Upogebia miyakei from Ryukyu Islands, Japan (Decapoda: Thalassinidea:
- Upogebiidae). J. Mar. Biol. Assoc. UK. 88: 125–131. (DOI: https://doi.org/10.1017/S0025315408000234)
- Itani, G. (2004) Distribution of intertidal upogebiid shrimp (Crustacea: Decapoda: Thalassinidea) in Japan. Contr. Boil. Lab. Kyoto Univ. 29: 383‒399.
- Komai, T. (2005) A new species of Upogebia (Crustacea: Decapoda: Thalassinidea: Upogebiidae) from Okinawa Island, Ryukyu Islands. Species Diversity 10: 259–268.
- Komai, T. (2020) Confirmation of the occurrence of a little known upogebiid mud shrimp Upogebia pugnax de man, 1905 (Decapoda: Gebiidae) in Japanese waters. Fauna Ryukyuana 53: 19-29.
- Futuyma, D.J. (1998) Evolutionary Biology. Third Edition. Sinauer Associates, Inc. Massachusetts.
- 池田嘉平・稲葉明彦(編)1971. 日本動物解剖図説。森北出版。
- Nanri, T., M. Fukushige, J.P. Ubaldo, B.-J. Kang, N. Masunari, Y. Takada, M. Hatakeyama, and M. Saigusa (2011) Occurrence of abnormal sexual dimorphic structures in the gonochoristic crustacean, Upogebia major (Thalassinidea: Decapoda), inhabiting mud tidal flats in Japan. J. Mar. Biol. Assoc. UK. 91: 1049‒1057. (DOI: https://doi.org/10.1017/S0025315410001712)
- Ngoc-Ho, N. (1994) Notes on some Indo-Pacific Upogebiidae with descriptions of four new species (Crustacea: Thalassinidea). Mem. Queensland Mus. 35: 193-216. (ISSN 0079-8835)
- 山田真弓・西田誠・丸山工作(1981)進化系統学。裳華房
- 西村三郎(1983)動物の起源論。中公新書。
- Saigusa, M., Y. Hirano, B. J. Kang, K. Sekiné, M. Hatakeyama, T. Nanri, M. Hamaguchi and N. Masunari (2018) Classification of the intertidal and estuarine upogebiid shrimps (Crustacea: Thalassinidea), and their settlement in the Ryukyu Islands, Japan. J. Mar. Biol. Oceanogr. 7:2. (DOI: 10.4172/2324‒8661.1000192
- リン・マルグリス, カーリーン・シュヴァルツ(1987)図説・生物界ガイド:五つの王国 。日経サイエンス社。
3.PDFファイルの請求
生物多様性研究・教育プロジェクト Research Reportsは,現在6つのジャンルに分けて出版されている。最初は,6つ目のジャンルとして「進化生物学博物館」を設けたが,第5報から名称を「比較系統学と進化」に変更した。比較系統学の分野では,論文を書く際にはMaterials and Methodsの単元に必要な事項を要約する必要がある。(分類学とは大きく異なる。)顕微鏡写真も多用され,図の中には形態の名称や器官の特徴を記入しなければならない。記事はHPで公開しているが, PDFファイルも同時に作成しており,各地の教育・研究施設に送付されている。
学術・教育目的で記事を利用するのであれば,公開しているメールアドレス(saigusa@biodiv-p.or.jp)に記事のタイトルを連絡いただければ,PDFファイルをお送りすることが可能である。ただ,公開しているメールアドレスは国内外からのジャンクメールでごった返しており,発掘するのに少し時間がかかるかもしれない。メールは,定期的にまとめて削除しているので,一緒に削除してしまったら申し訳ない。
なお,分類学の場合には,GoogleやGoogle ScholarからPDFファイルを入手することは難しい文献が多い。申し訳ないが,生物多様性研究・教育プロジェクトでは文献入手のサービスは行っていない。