
2024年7月22日(月)
1.はじめに
「泥」(mud)と「砂」(sand)は,粒子の大きさが異なる。泥は,粒径が62μm以下の微粒子のことを言う。詳しくは,粒径が4μm以下をクレイ(clay),4μmから62μmまではシルト(silt)と呼んでいる。一方,砂は泥よりも粒が荒く,粒径は62μmから2mmの範囲の粒子のことを言う。泥は陸上起源である。主には泥岩(mudstone)が風化して生じるのだろう。一方,砂の起源は陸上と海洋の両方だと思われる。陸上では砂岩や石灰岩が風化して生じるのに対し,海洋ではサンゴ塊(石灰岩)が波による破砕を受けて生じたものだろう。なお,粒径が2mm以上になると砂利(gravel)と呼んでいる。
河川の海への出口にあたる河口(estuary)には,陸上から運ばれてきた泥,砂,砂利,岩石が堆積する。河川の勢いが強ければ,これらは河口を越えて海洋のはるか遠くまで運ばれ,海底に堆積する。海洋起源のサンゴ塊は,荒い波に砕かれて,海岸や海底に堆積することが多いだろう。日本列島であれば,波の荒い太平洋側の海岸には砂が堆積し,潮位の変動が少ない日本海側の海岸,瀬戸内海のような内湾,大きな河川の河口には泥が堆積することが多い。おそらく最近(1,000年前とか2,000年前)のことだろうが,多くの内湾にはクレイを多く含む泥(田んぼの泥と同じ)が大量に堆積し,数メートルにもおよぶ厚さの軟泥層(ヘドロとも言う)が形成されている海岸もある(有明海や笠岡湾)。
生態学的に見た場合には,泥と砂では大きな違いがある。泥の層は有機物を多く含むのに対し,砂の層には少ない。だから砂質の干潟にすむ生物(例えばアナエビ類)は,ろ過摂食(filter feeding)が成り立ちにくい。一方,アナジャコや多くのスナモグリのように,泥の層に住む生物はろ過摂食をしている。しかし,シルトやクレイでできた層は水はけが悪い。水はけが悪ければ,土壌の中に酸素が供給されなくなる。土壌の有機物は嫌気性のバクテリアによって分解されるため,分解産物として硫化物が生成される。土壌の色は黒くなり,しばしば悪臭を伴う。
新生代における陸上植物の繁栄によって,地球環境は大きく変化した。特に海洋では,陸上から泥とともに大量の有機物が流入し,河口や海岸の著しい富栄養化(eutrophication)を引き起こしている。無機物にまで分解できなかった有機物は,植物によって粉砕された微小粒子(シルトやクレイ)に混入し,海岸や河口域に軟泥層(いわゆるヘドロ)として堆積している。軟泥層の堆積は急速な勢いで拡大している。軟泥層には酸素が行き渡らないため,ヘドロの堆積は生物種(主にはベントス)の顕著な減少,つまり深刻な海洋汚染を引き起こす。海を汚しているのは人間だけではない。陸上における被子植物の多様化も海洋汚染に立派に貢献している。
新生代第四紀に急速に広がった海洋汚染にうまく適応した生物がいる。それがアナジャコやスナモグリの仲間である。分類学上はエビやカニと同じく十脚甲殻類(decapod crustacean)に属する。どちらも奇妙な形態をしている。アナジャコでは,目(眼柄)は,頭部先端の額角(rostrum)の下に収納されている。額角の先端には複数の堅固な角(棘というよりも小さな角という感じ)が配置され,巣穴への侵入者には角を突き出して外に追い払っている。鋏脚(第1歩脚)の掌節(propodus)に多数のトゲを備えた種類もいる。アナジャコ類は,肉眼で見る限りはみなよく似ているが,双眼実体顕微鏡(binocular microscope)の下で観察すると,特に体の前半部の形態は著しく多様化している。あえて例えると,体の前半部(頭部から胸部にかけて)はホ乳類のサイのように鎧兜(よろいかぶと)を身にまとった堅固な構造をしているが,胸部から後ろ(腹部と尾部)はゴカイやミミズ(環形動物)のように柔らかい構造(一応十脚甲殻類の構造を維持しているので,そこまで柔らかくはないか・・・)になっている。泥の中に巣穴を作って住むという生態学的な制約もあって,どの種類も体はみな細長い。
アナジャコの住む泥干潟の環境は一様ではない。底質(substrate)は軟泥層の下に砂の層がある海岸や河口がある。サンゴ塊の残骸が埋まっていたり,大小さまざまな径の砂利がたくさん埋まっていることが多い。海岸によっては,泥岩の中に空洞がたくさんあって,多数の捕食者が住んでいそうなハビタットがある。そういうところでは,巣穴は縦方向ではなく水平方向に作られ,捕食者に遭遇した時にはサーベルのようなトゲで追い払うか,複雑な通路をすばやく逃げ去るのだろう。巣穴を作る底質の条件や強力な捕食者の有無によって,どんな形態(特に頭部と鋏脚)をもった個体が生き残りやすいか,大きく異なる可能性がある。底質(substrate)の構造や硬さ,そして捕食者の有無がアナジャコ類の形態の多様化をもたらしているようだ。
現在私が研究しているのは,河口域の砂利に富んだ泥干潟に生息するアナジャコ(gebiidean decapods)である。和名はヨコヤアナジャコ,学名はUpogebia yokoyai(リンネの2命名法に基づく表記)である。日本の固有種とされているが,どうしてそう断言できるのか?
その疑問に答える前に,ヨコヤアナジャコと認定されている種(species)は本当に1種類なのか,まずその疑問に答える必要があると思われた。
<ヨコヤアナジャコ研究の前段階>
ヨコヤアナジャコ(Upogebia yokoyai)は,琉球弧(Ryukyu Arc)を含む日本列島の沿岸域に生息している。伊谷行氏(2004)によれば,ヨコヤアナジャコは日本の固有種で,東北地方から南西諸島の内湾や河口の干潟に生息しているとのことである。
どういうきっかけでそうなったのか全く覚えていないが,ヨコヤアナジャコは小さな個体が生息している泥干潟と大きな個体が生息している泥干潟があり,2つの個体群の間でどうしてこんなに体長や体重に差があるのかを調べてみようということになった。
この事例の場合には,学生の基礎学力が不足していたために研究指導が難しく,研究から参考論文(博士号取得の審査に必要)の作成まで全部私が行う事態になった。しかもそんな学生が複数いたので,そのころは大変な毎日が続いた。
私に対して,全然専門的なことがわかっていないと真剣に怒る学生(複数)がいた。言い訳ではないが,教員は多くの学生の研究指導を統括する義務を負っている。学生が1人2人なら問題はないが,博士課程(前期と後期)の学生の中には,基礎学力が不足して自分では研究を進められない者がいる。そういう者に対しては,私が代わって研究を行い,論文(参考論文)の作成を行う羽目になる。(放っておくと,内部告発で不適切な研究指導を認定され,処分が下る。)そんな学生が入ってくると,研究指導に大半の時間を使うことになって,他の学生には十分に目が行き届かなくなる。そうすると,あの教員は指導能力がない,などと各所に触れ回る学生が出てくる。
アナジャコの研究を始めたのが2004年あたりで,2009年からはブッポウソウの研究が重なった。教育活動の激務もあって,2015年までの6~7年間は,毎日がまさに地獄の日々であった。しかし,逃げたら敵の思うツボなので,それは絶対にしなかった。同僚の教員にはよほど嫌われていたのだろう。スーパーで私の姿を見てさっと逃げてゆくやつもいた。
<ヨコヤアナジャコの分類に関する新しい問題>
ヨコヤアナジャコは,広島県の廿日市市(はつかいちし)にある佐方川には大型の個体が住んでおり,高知県の甲殿川(こうどのがわ)には小型の個体が住んでいることに注目し,それぞれの個体群の生活史(life history)を調べてみた。大潮の干潮時の関係で,広島は日帰りが可能だが,高知は一泊して岡山に帰ったと思う。結果は,生活史のパターンは両者の個体群で大きく異なっていた。論文を出した時点では,2つの個体群の生活史の違いは,互いに別な種ではなく,佐方川と甲殿川の富栄養化の違いに起因すると考えていた。
これもきっかけは思い出せないが,当時博士後期課程に在籍した姜 奉廷さん(発音はカン・ボンチョン?)に遺伝子解析と分子系統図の作成をお願いした。姜さんは,立正大学の関根一希さんと共同研究を行って,日本列島の河口に分布するヨコヤアナジャコの遺伝子型を調べた。ヨコヤアナジャコの採集は,瀬戸内海(本州沿岸と四国沿岸),日本海沿岸,琉球弧は私が中心となって行い,九州は南里敬弘氏(今どこで何をしているか知らない)が中心となって行った。
姜さんと関根さんの分析の結果,日本列島に分布するヨコヤアナジャコは,遺伝子型から見るとAからFの6つのクレード(clade)からなっていることがわかった。クレードA, E, Fは,本州,四国,九州の沿岸で見られ,B, C, Dの遺伝子型は,琉球弧で見られた。
佐方川の個体群は,瀬戸内海,日本海側, 九州に広く分布するクレードAに,甲殿川の個体群は太平洋岸に分布するクレードEに属していた。全部で6つの遺伝子型のうち,佐方川と甲殿川に生息する個体群は,生活史のパターンや成長度の違いから互いに別種と考えられることに気づいた。同じ生息場所から同時に2つの遺伝子型が検出されることがあり,形態は酷似しているが,同一の干潟に別の種が同居しているのではないかと推察された。
分子系統解析の問題点は,形態との関連に言及することなく,研究は遺伝子型から見るとこう考えられるという所で終わっていることだ。分子系統学の研究者は遺伝子解析には夢中になるが,形態には無関心の人が多いのだろう。逆に,分類学者は分子系統学を理解していない。依然として平安時代の貴族のカルタ遊びみたいなことにこだわっている。要するに,どちらの学者も専門性を追求するあまり,著しく視野が狭くなっているのだと思う。
遺伝子解析から2つのクレードが検出されれば,形態的にも違いが出るはずである。双眼実体顕微鏡下でよく調べれば,明確な違いを見つけることができるはずだ。私はいつも両方の視点で考えるので,どちら側の学者からも毛嫌いされるのは承知している。
琉球弧に分布するB, C, D型については生活史を調べることは難しいが,形態の違いがあるはずと考えた。採集は,石垣島,西表島,宮古島,そして沖縄本島の海岸や河口の干潟で行われた。私はもともとフィールド・ワークが大好きである。琉球弧には,2024年6月中旬から7月上旬にかけて,私一人で行った。西表島と沖縄本島には,琉球大学の熱帯生物圏研究センター(西表研究施設と瀬底研究施設)があって,そこに宿泊して採集ができる。どちらも快適な生活を送ることができた。宮古島では,伊良部島にある民宿「まるよし」に泊まった。民宿と言ってもビジネスホテル形式で,管理人のおばさん,おじさんもとても親切だった。
ヨコヤアナジャコの生息場所(ヤエヤマヒルギの泥干潟)は,「まるよし」から車で5分のところにあった。生息環境がよく似ているので,ここに住んでいるのはヨコヤアナジャコ(Upogebia yokoyai)に間違いないと思っていたが,双眼実体顕微鏡で調べてみると,全然違った種類だった。駒井智幸氏の言うUpogebia pugnaxか,それに酷似した別種だろう。形態だけでなく,生息場所の環境特性,体色,体の大きさ,分子系統解析の結果を総合的に考慮して種を認定したい。液浸標本の形態だけで種を判定することは,私にはできない。
琉球弧で最初に行ったのは石垣島である。インターネットで予約して止まった民宿(?)は,どっかの大学の寮みたいな古い木造のアパートだった。部屋にはエアコンと机と布団があるだけ。畳の上には恐ろしい数の微小なアリが矢のように走り回っていた。木造アパートは多くの無脊椎動物に占領されていた。一泊3,000円だったが,こういうところは私のような年寄りには向いていない。若い人向けのアパートである。石垣島では安くて快適な宿泊場所が見つかっていない。
2.ヨコヤアナジャコ(Upogebia yokoyai)に関する基本情報
<Upogebiaの語源>
Upogebiaという属は,1814年にLeachという人が命名した。語源については不明である。エジプト神話にGebという大地の神(男神)が出てくる。Gebiaはその大地の神が住む場所か,大地の神の一族という意味があるのかもしれない。一方,Upoはタガログ語で「ひょうたん」を意味する。つまりUpogebiaは,「ひょうたん型をした大地の神」という意味なのではないかと推察した。
しかし,なぜ「ひょうたん」なのか? そう思って頭胸部の背面を双眼実体顕微鏡で観察すると,アナジャコやヨコヤアナジャコの場合には,確かにひょうたん型をした溝(groove)があるのがわかる。この溝は多くの十脚甲殻類にみられるが,ひょうたん型は一部のアナジャコに限られるように思われる。
<ひょうたん型の溝の起源>
ひょうたん型の溝(groove)は,アナジャコ,スナモグリ,一部のヤドカリ,オキナワアナジャコ,そしてアナエビ類にみられる構造である。タラシナ線と呼ぶ形態学者もいるが,私は使用していない。ひょうたん型の溝(模様)の起源についての説明には,節足動物の発生学に関する知識が必要である。しかし,日本の節足動物(系統)発生学者,例えば椎野季雄氏(故人)や関口晃一氏(故人)に尋ねても,納得できる答えは出してくれないだろう。Glaessner (1969)が中生代の化石のNephropsやHomarus(Astacidea)の胸部背面から胸部側面にみられるいくつかの溝について言及しているが,進化した十脚甲殻類の胸部背面にみられる「ひょうたん溝」については触れていない。
ひょうたん型の形態は,十脚甲殻類の進化と密接にかかわっている。ひょうたん型の溝の起源は,第3顎脚(third maxilliped)が生じる体節(第3体節?)と第1歩脚(鋏脚)の生じる体節(第4体節?)の「くびれ」である。十脚甲殻類の場合には,第1顎脚から第3顎脚が口器(こうき)の周辺に圧縮されてしまった結果,このくびれの行き場がなくなって頭胸部の背面にひょうたん模様として残ったものだろう。だから,この溝がどんな形状をしているかは,アナジャコ類の系統を強く反映していると考えてよいだろう。
なお,ひょうたん型の溝は同じ十脚甲殻類のヤドカリ類には残っているが,カニ類では消失している。ひょうたん溝の形態(morphology)が十脚甲殻類の進化にともなって変形する過程については,別の記事で議論したい。
節足動物の系統発生学から提案される形質(ひょうたん溝の形状)を基礎に置けば,アナジャコ類をいくつかの属(genus)レベルでまとめ上げることができると思う。酒井勝司氏の分類は,啓示を基礎とする宗教者のやり方と同じである。分類学を自然科学として世に広めようと思うならば,判断する客観的「基準」(standard),しかも現代的基準を明確にした方が理解を得られやすいと思う。
私は酒井勝司氏の分類学は理解できない。当時博士後期課程に在籍した平野優理子さんは,酒井勝司氏に教えを受けた。しばらくして酒井氏の学問には大きな問題(questionではなく,problems)があることが分かった。酒井氏の学問は古すぎるのである。平野さんはそこをよくフォローしながら, 博士論文を完成させた。その意味では平野さんは非常に優秀な学生だったと思う。(平野さんの欠点は,何か尋ねるとすぐ怒りだすこと・・・。たくさん怒られた記憶がある。)
DworschakやNgoc-Hoなども酒井氏とよく似た古い学者だと思う。人間は,今までに培った自己の権威(既得権ともいえる)を守りたいばかりに,自分に疑義を唱える者を徹底的に排除することがある。上記の3人には,そんな共通した特徴がある。
酒井氏は人格的にも大きな問題を抱えた人で,研究室は大きな被害を被った。(他の施設でも同様な話を聞いている。)・・・が,人格的な問題は横に置くとして,このままアナジャコ類の研究を終わらせては,私の研究指導の無能さ・無責任さが残るだけになる。平野さんに変わって,私が分類学の分野に参入することになった。平野さんとはもう交流はない。分類学の権威の方々にとっては,大変迷惑なことは重々承知している。
<アナジャコ類の起源と進化>
アナジャコの起源はサンゴ礁原の干潟に生息するアナエビ(Axiidea)の1種だろう。ただし単系統(monophyly)か多系統(polyphyly)かを判断する証拠はまだない。琉球弧に分布するヤハズアナエビは,生息環境から見てアナジャコの祖先に近いような気がする。しかしKensleyやPooreらの論文を見ると,ヤハズアナエビよりはもっと近い種がいそうである。一方,アナエビ類の起源は,中生代の三畳紀に砂泥底の内湾に生息したロブスター(Archaeohomarus:仮称)であろう。
アナジャコをmud shrimpと呼ぶ人は多いが,shrimpはコエビの仲間を指す。もちろんmud lobsterもよくない。アナジャコはもうロブスターではないのだから・・・。学名にgebiaという名称が使われているので,私は現在gebiidean decapodという名称を使っている。Gebian decapodでもOK。
スナモグリも,遠い祖先はロブスターだろう。中生代の三畳紀末にサンゴ礁原に進出したロブスターが,直接スナモグリの祖先に移行したのかもしれない。スナモグリはghost shrimpと書く人が多いが,アナジャコと同様にshrimpではない。ロブスターでもない。Callianidean decapodとか,callianassidian decapodという表現の方が実態に近い。しかし,「それに従わねばならない」と言っている訳ではない。
<アナジャコ類の研究を許可したauthority>
最近は上記の項目について明記させるjournalが増えている。私の場合には,独立性(independency)が強く出てしまうので,権威の筋からは強いお怒りの言葉を頂戴することになるだろう。・・・が,そんな非難に屈するほど暇ではない。村八分になれば,自分の好きなことを自由にやれるだけのこと。内容については,ブッポウソウの研究の記事に記したが,ナンバーは忘れてしまった。
<分類(classification)に関して権威者に尋ねることはない理由>
生物の種名を決定するのは分類の権威者という習慣が根付いている。しかし,権威者の視野は一般に非常に狭く,しかも神がかった判断をする人が多い。そういう学者に客観的な証拠を突き付けても理解を得られる可能性は低い。私は,権威のある学者の言うことはいつも正しいとは思っていない。同じ人間であり,どんなに偉い人でも間違い,誤解,判断ミスはたくさんある。特に,分類学のように主観的な証拠の上に立って種を判断すると間違いが起きやすい。
生態学も同じ。統計的検定を使って検証すること(verification)は現代的手法として認められるが,出発点の「仮定」あるいは「基準」に難がある。仮定が事実や確かな基準の上に置かれず,(神の)啓示に近いところに置かれると,非現実的な話ができあがる。要するに,生態学は自然科学ではなく,社会学や経済学と同じ文科系の学問である。伝統的な分類学も,判断する客観的基準を明確にしないと,宗教学と似た学問になる。これも文科系の学問だろう。
高等学校の生物の教科書は,生物学の基礎を学ぶという観点から,実証的な側面を重視している。生態学の分野では,仮説を立てて統計的手法を用いて「検証」したと言っても,仮説を「実証」する(prove)訳では無い。だから,実証的志向の生物の教科書では,生態学や古典分類学は身の置き場に困るだろう。生態学は,今流行りの「データサイエンス」に身を置いたらうまく収まるように思える。
分子系統学の分野では,遺伝的な違いを定量的に検出することができるが,ダーウィン,ヘッケル,木村資生がやったような実証的進化論(どれも仮説だが・・・)まで提唱する人は少ない。私は,間違ってもいいから,権威者の仮説を鵜呑みにせず(もちろん理解しようと努める),(決して優れてはいない)自分の頭で思い切り考えて話(仮説)を作ってゆきたい。だから,ゴールは生態学や分類学と同じなのだろう。野鳥の会の何某のように,私の記事は一切見ないと公言している者がいるが,そういうことで解決できる問題ではない。
<アナジャコ類の日本列島への定着>
アナジャコ類の祖先は,サンゴ礁原にできた砂干潟(sand tidal-flat)に住んでいたアナエビ(Axiidea)の仲間であろう。アナジャコの生息場所を調べると,軟泥層の下には砂利や死んだサンゴ礫(れき)(rubble)が出てくることが多い。おそらく地形の変動によって,アナエビの生息場所である砂干潟に泥が堆積するようになったのだろう。アナエビはまず泥の層に潜り,その下のサンゴ礫に作られた巣穴で生活するようになったのではないだろうか?
摂食様式も,サンゴ礁原に生えた藻類やアマモを切り取って食べる性質から,巣穴に流入する泥の微粒子を口器(第1顎脚,第2顎脚,第3顎脚)と歩脚の腕節(carpus)や長節(merus)に生えた無数の長い毛(setae)で濾しとって食べるデトリタス・フィーディングに変わっていったと思われる。生活の変化,食性の変化の過程で,形態形成(metamorphosis)に起きた微小な突然変異が蓄積され,典型的なアナジャコのフォルムができたのだろう。ヨコヤアナジャコのひょうたん溝も,頭部の形態変化と前後して生じた(改変された)と思われる。
アナエビからアナジャコへの移行は徐々に起きていると考えられるが,これはアナジャコだというフォルムが完成した時代はまだよくわかっていない。インド洋と西太平洋地域において,泥岩の地層が隆起して大洋島や大陸島の形成が活発になった時期があるのだろう。台湾もその時代に出現したと推察される。河川によって運ばれる泥は,被子植物が起源であり,有機物を多く含む土壌だったろう。島嶼(とうしょ)地域では河口域に大量に堆積する時代が訪れたのは,私の予想だと新生代第四紀(258万8000年前から現在まで)ではないかと思う。つまり,200万年前ごろから,地球の河川(河口)の環境は大きく変化し,その変化と前後してアナジャコが誕生したのではないかと思われる。ちなみに,西表島が島として海上に姿を現したのは1,300万年前(中新世),宮古島は200万年前(更新世),本州や九州,四国は2,000万年前(中新世)だろう。
琉球列島の島々の海岸や河口の泥岩露頭には,何種類かのイシアナジャコが住んでいる。それぞれの種の分岐年代を推定すると,古いもので50万年前,新しいもので20万年前ぐらいだったかと記憶している。種分化(speciation)が起きた場所は不明だが,赤道から台湾の間に点在する大小多くの島々の河口であろう。
地球の自転により,海洋にはコリオリの力が働いている。ユーラシア大陸の東岸だと,北半球の赤道付近では西向きの海流が生じる。この海流(黒潮)は,フィリッピンの東側で北に向きを変え,台湾の近海から琉球列島を通って四国沿岸に達する。アナジャコは,そのハビタットで世代を重ねるうちに,海に流れ出た幼生(larva)が黒潮に乗って,日本列島の島々に流れ着いて定着したのであろう。イシアナジャコ類と比べて,ヨコヤアナジャコは種(今はクレードの段階に身を置いている)間の形態の違いが小さい。だから,イシアナジャコ類よりも少し遅れて種分化(speciation)が始まった可能性が高い。
クレードAが一番古く,30万年前ぐらいだろうか?続いてEとF。20万年から30万年前あたりと想像する。琉球列島に分布するB, C, Dは,10万年から20万年前と予想しているが,大外れということもありうる。大きな問題は,種分化(生殖隔離)がどこで起きたのかということだ。
10万年前とか20万年前だと,日本列島にはまだヒルギ林(マングローブ)はなかっただろう。つまりヨコヤアナジャコの生息に適する環境(受け入れ環境)はまだ整っていなかった可能性が高い。もしそうだった場合には,ヨコヤアナジャコの原種が日本列島に流れ着いて,ほんのわずかな間(例えば2,000年とか8,000年とか)に,それぞれの島で微小な形態の違いが生じたという仮説になる。この仮説を認めるならば,例えば南西諸島に分布するUpogebia pugnaxに,原種(ニューカレドニア?)と同じ種名を付与するのは合理的判断と言える。Upogebia saigusaiなどは絶対あり得ない話になる。
しかし,私はこの仮説の信ぴょう性は低いと思う。なぜ低いかについては,近いうちに客観的な証拠を集めて論じてみたい。
私が別種とみなした種類に関して,その判断が正しいかどうかをどこかの権威に尋ねることはしない。私のやっていることは,できる限り多くの客観的事実(証拠)をもとに,どういうことが考えられるかという仮説(hypothesis)をパブリッシュすることである。自分の意見をパブリッシュすると言い換えてもよい。私の言っていることが権威者に認められたから論文をパブリッシュできるとか,認められなければパブリッシュできないとか,そういう考えで生きている訳では無い。権威あるreviewerの中には,異常なほどの偏見をお持ちの方がたくさんいるのではないか?偏見のある方々のご機嫌を伺って有名なjournalに論文を出すことができても全然嬉しくない。
<アナジャコ類の分類には,形態的特徴だけでなく,体色,体の大きさ,生態に関する情報が不可欠>
形態の微小な違いをめぐって,形態情報(スケッチ)だけでどの種と認定するかで争うのは止めにしたい。それよりは研究の再現性を考慮し,形態学的な記載に加え,採集場所の位置と生態学的な環境を正確に記載することによって,種を分ける方が現実的である。特にアナジャコに関しては,環境条件が少しでも違えば,それが種の違いに反映されるように思う。間違っていると指摘されれば,同じところに採集に行って研究をやり直すことができる。生息場所の位置が共有されれば,研究者の間に,あそこに住んでいるのはあの種の可能性が高いというコンセンサスができる。
アナジャコ類の場合には,種の判定(identification)に形態ばかりではなく,集団遺伝学的思考,分子系統解析,環境条件に関する知見も導入できればメリットは大きい。しかし,そうなると論文をアクセプトしてくれるjournalの数は一気に減少する。集団遺伝学における遺伝子浮動のプロセス(生存に有利な突然変異でも,個体群の中に定着するには時間がかかる)と同様に,しばらくは不利な状況を耐え忍ぶ必要があるだろう。
<何をパブリッシュするか?>
少なくとも私は,権威者の発する直感的な啓示をありがたく思い,神の啓示によって自分自身の人生を作るのではなく,できるだけ多くの客観的証拠に基づいて自分自身の頭で考えて最終的な判断を下すという道を選んできた。要するに,生態学者が大変嫌う作業であるdescriptiveなデータを仮説とともにパブリッシュしたいのである。descriptiveなデータがないと,どんな物語も可能になる。
ラジオやテレビでサプリや薬の効果を懸命に説いている者をよく見かける。怪しげな根拠を持ち出して,特定のサプリや薬の有効性を鬼の首を取ったように自信満々に話をしているのを見ると,こいつ頭がおかしいのでは・・・と思ったりする。「進撃の巨人」も世に大受けした。アニメと同じ調子で自然科学の世界に身を置く人たちが増加しているのかもしれない。実証的科学者にも根はそんな人も多い。
3.採集,撮影,原稿執筆の基本情報
<採集> 三枝誠行(NPO法人 生物多様性研究・教育プロジェクト,常任理事)。
<撮影者> 三枝誠行
<撮影機材> ボディはEOS 7D(Canon)。これに CANON Compact-Macro Lens EF 50mm 1:2.5(接写レンズ)をつけた。これは非常に有効だが,泥がいっぱい着くので海岸に持ってゆくのは危険。もう一つはEOS 7DにTAMRON 28‒300mm F/3.4‒6.3 Di VC PZD(ズームレンズ)を装着した。これも風景と昆虫撮影には役立つ。顕微鏡撮影装置も持って行ったが,あまり役立たず,途中で送り返した。その他,SONYのビデオカメラ(HDR‒CX 680)も使ったが,風景と昆虫には使えるが,海に持ってゆくことはできなかった。NIKON CoolpixやRICOH WG‒50も使ってみたが,あまりきれいには写らなかった。
<執筆> 三枝誠行・増成伸文(NPO法人 生物多様性研究・教育プロジェクト)。
4.参考文献
・Castro, P., and M.E. Huber (2005) Marine Biology, Fifth Edition. McGrowHill Higher Education, Boston.
・藤本 潔・三浦正史・春山成子(2015)西表島仲間川低地におけるマングローブ林の立地形成過程と地盤運動。Mangrove Science 9: 3‒15.
広島大学生物学会(編)1971. 日本動物解剖図説。森北出版。
・Itani, G. (2004) Distribution of intertidal upogebiid shrimp (Crustacea: Decapoda: Thalassinidea) in Japan. Contr. Boil. Lab. Kyoto Univ. 29: 383‒399.
・Hirano, Y., and M. Saigusa (2008) Description of the male of Upogebia miyakei from Ryukyu Islands, Japan (Decapoda: Thalassinidea: Upogebiidae). J. Mar. Biol. Assoc. UK. 88: 125–131. (DOI: https://doi.org/10.1017/S0025315408000234)
・木村政昭(1996)琉球弧の第四紀古地理。地学雑誌105: 259-285.
・木村資生(1988)生物進化を考える。岩波新書。
・小竹信宏・三浦正史・奈良正和(2013)沖縄県西表島の中部中新統西表層最上部の地質年代と堆積環境。地質学雑誌 119: 701‒713.
・Nanri, T., M. Fukushige, J.P. Ubaldo, B.-J. Kang, N. Masunari, Y. Takada, M. Hatakeyama, and M. Saigusa (2011) Occurrence of abnormal sexual dimorphic structures in the gonochoristic crustacean, Upogebia major (Thalassinidea: Decapoda), inhabiting mud tidal flats in Japan. J. Mar. Biol. Assoc. UK. 91: 1049‒1057. (DOI: https://doi.org/10.1017/S0025315410001712)
・Nybakken, J.W. (2001) Marine Biology: An Ecological Approach, Fifth Edition. Benjamin Cummings, San Francisco.
・Saigusa, M., Y. Hirano, B. J. Kang, K. Sekiné, M. Hatakeyama, T. Nanri, M. Hamaguchi and N. Masunari (2018) Classification of the intertidal and estuarine upogebiid shrimps (Crustacea: Thalassinidea), and their settlement in the Ryukyu Islands, Japan. J. Mar. Biol. Oceanogr. 7:2. (DOI: 10.4172/2324‒8661.1000192
・椎野季雄(1964)動物系統分類学 7(上)節足動物(Ⅰ)総説・甲殻類。中山書店。
・Ubaldo, J.P., T. Nanri, Y. Takada, and M. Saigusa (2014) Prevalence and patterns of infection by the epicaridean parasite, Gyge ovalis and the emergence of intersex in the estuarine mud shrimp, Upogebia major. J. Mar. Biol. Assoc. UK. 94: 557‒566. (DOI: https://doi.org/10.1017/S0025315413001392)
・Williams, A.B. (1995) Taxonomy and Evolution. In: J.R. Factor (ed.) Biology of the Lobster Homarus americanus. Academic Press, San Diego.
・Yamasaki, M., T. Nanri, S. Taguchi, Y. Takada, and M. Saigusa (2010) Latitudinal and local variations of the life history characteristics of the thalassinidean decapod, Upogebia yokoyai: A hypothesis based on trophic conditions. Est. Coast. Shelf Sci. 87: 346‒356. (https://doi.org/10.1016/j.ecss.2010.01.017)
・山田真弓・西田誠・丸山工作(1981)進化系統学。裳華房。
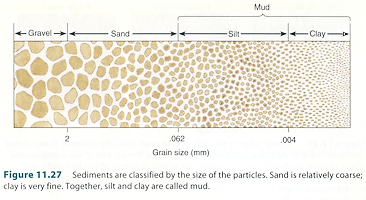
図 1.粒径(grain diameter)を基礎とした泥(mud)の定義。
図 1 は,Castro and Huber (2005) Marine Biology から転載した。
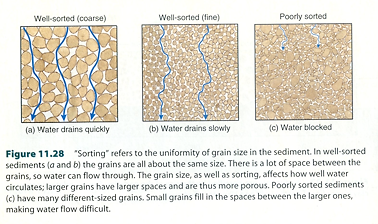
図 2.泥の堆積層における粒度(grain size)と水の透過性の関係。
(Castro and Huber (2005) Marine Biology から転載)
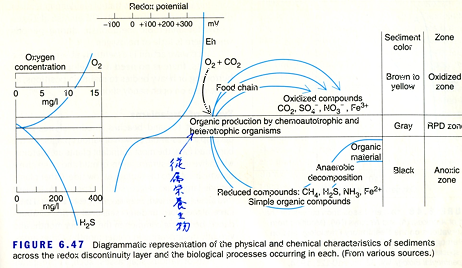
図 3.堆積した泥に付着している有機物の細菌(bacteria)による分解。酸素が行きわたっている層では,好気性細菌によって分解され泥の色は茶色から黄色になる。酸素が供給されない層で分解が行われた場合には,分解産物として硫化物,アンモニア,還元化した鉄が生じ,層の色は黒色になる。場所によっては,悪臭を伴う。図 3 は Nibakken (2001) Marine Biology から転載。

図 4.川平湾に流れ込む小さな川の河口(干潮時)。写真左側はヤエヤマヒルギ,手前にはモクマオウの枝が写っている。ユゴイの群れが泳いでいた。

図 5.川底に作られたヨコヤアナジャコの巣穴。穴の深さは 20~30cm ぐらいのものだろう。川の深さはせいぜい 20cm 程度なので,長靴で歩ける。アナジャコはスコップを使えば簡単に採集できそうである。この小さな川は,川平湾に流れ込む。2009 年に,姜さんが川平で採集されたヨコヤアナジャコのDNA のシークエンスを行った。その時には,石垣島のヨコヤアナジャコは Clade B に分類された。2024 年に採集に行くまでは,石垣島のヨコヤアナジャコは,石垣固有種と思っていたが,沖縄本島にも分布している可能性が高い。西表島にも分布する可能性が高い。

図 6.名蔵小橋(図 7)の下を流れる川(干潮時)。この川では時々河口閉塞が起きるかもしれない。モクマオウがヤエヤマヒルギと同じ位置に張り出しているので,塩分濃度が低いかもしれない。川底にはヨコヤアナジャコの巣穴は見つからなかった。
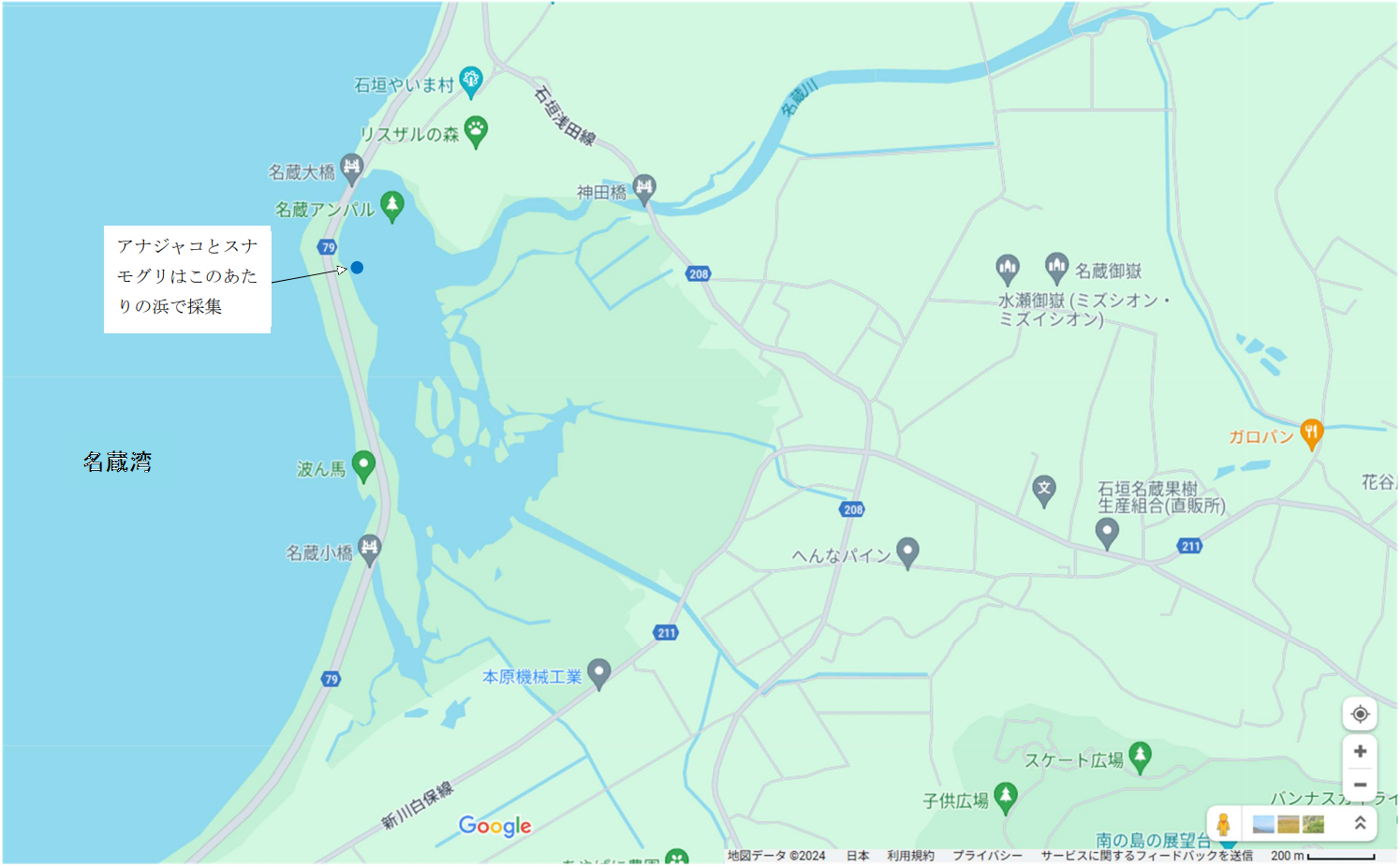
図 7.名蔵川の河口におけるアナジャコとスナモグリの採集。河口域は広いが,海水の出入りする名蔵大橋の下が狭いので,河口の奥(上流域)は塩分濃度が低いかもしれない。名蔵大橋の近くの浜では,アナジャコとスナモグリを採集することができた。川の近くの泥干潟(干潮時)でいくら頑張っても全然採集できなかったので,疲れてしまって上流に行くのは止めた。

図 8.干潮時に名蔵川河口に現れた泥干潟(名蔵大橋から 150m ほど上流)。干潟の表面には砂が堆積している。川に近づくにつれ泥の割合が増加するので,川底は柔らかい土壌になる。潮が一番引いたころには,向こう岸(200m ぐらいあるか?)まで長靴で歩いて行ける。

図 9.泥干潟に作られたヨコヤアナジャコ(?)の巣穴。アナジャコの巣穴は,穴の表面が粘土(clay)でコーティングされている。生息場所の底質は波で細かく砕かれたサンゴ塊や貝殻だろう。粒径の細かい泥は,還元化して黒くなっている。

図 10.遺伝子解析の結果から,石垣島には Clade B のみが分布しているが,これが B タイプなのか検討中である。体長 6 ㎝は大きいタイプの種類である。頭部背面にはひょうたん模様があるので,ヨコヤアナジャコの仲間には間違いないが,本州,四国,九州に生息する個体とは,体色や体の大きさに違いがあるように見受けられる。細かい外部形態の違いから,別種と判断してよいか検討中である。自称専門家の方々にお伺いを立てたら,すべてヨコヤアナジャコとして処理されるのは間違いない。例えば,酒井勝司氏にお伺いを立てたら,私たちのアナジャコの研究は完璧にぶち壊されてしまう。
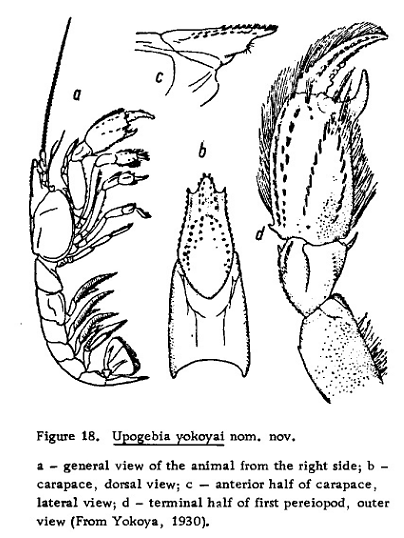
図 11.ヨコヤ氏が描いたヨコヤアナジャコのスケッチ。
大変失礼だが,このような図ではアナジャコの仲間であることはわかるが,属(genus)や種(species)になると全く判断できない。昆虫採集のように,野山を駆け巡って採集した後に図鑑を使って種類を調べるというような場合にはいいかもしれないが,専門的研究でこんな粗雑な情報が種名判断の基礎となっているのでは,分類学という学問そのものの意義が疑われる。リンネやラマルクの時代から何にも進歩していないではないか・・・。
論文がパブリッシュされたのは 1930 年なのだが,ヨコヤアナジャコに関してはそれ以来スケッチの報告はないかもしれない。だから,多くの専門家はこのスケッチをもとにヨコヤアナジャコを認定していたのだろう。繰り返すが,こんなスケッチでヨコヤアナジャコを認定するのは,宗教家にしかできないだろう。
こんな状況だから,スケッチは書かせてもらえるけれど,種名の方は自称専門家にお伺いを立てて判断してもらう習慣が根付いてしまったと思われる。ギルドと同じく,権威者がウンと言わなければ論文など出すことができない時代が長く続いた。伊藤篤太郎を破門にした矢田部良吉なども,その時代の典型的な人物だっただろう。
権威者のご機嫌取りをしないと研究をさせてもらえない分野は,なにも分類学に限らず,1960 年代から始まった微細構造形態学(透過型の電顕を使う),電気生理学,ショウジョウバエの遺伝学も同様であった。こういう権威主義的な生物学は,今はみんなつぶれてしまって,跡形も残っていない。十脚甲殻類の権威主義的分類学も,現代生物学の進歩に見放され,今や風前の灯火になっている。

図 12.名蔵川河口の中央部(干潮時)。泥干潟の所々軟泥層が堆積している。中央部にもアナジャコかスナモグリの巣穴がたくさんあって,中にいることはわかるが,川底を何度掘っても採集できなかった。巣穴は,20cm から 30 ㎝の深さまでは縦穴で,それから横穴になっているかもしれない。

図 13.名蔵川の上空に広がった入道雲。午後になると黒い入道雲が海の方(西)から現れた。名蔵湾の上空では雷が発生し,稲光がしてから雷鳴がとどろいた。黒い雲が干潟の上空に達すると雷が怖いので,一時土手に避難してやり過ごした。採集はいつも入道雲の流れる方向を見ながら行った。

図 14.干潟の上の水たまり。アナジャコやスナモグリを掘った後が水たまりとして残っている。半分は私が掘った穴だが,あとの半分は釣り餌として使うため,地元の人が掘ったと思われる。いずれにしてもこの干潟での採集は難儀した。石垣島ではヨコヤアナジャコの採集は難しい。

図 15.スナモグリ(種名は不明)。頭胸部の背面の模様(溝)はひょうたん型ではない。頭胸部背面の模様と額角(rostrum),および側方額角(lateral rostrum)をアナエビ(例えばヤハズアナエビ)と比較すると,多くの共通点を見出すことができる。

図 16.名蔵川の泥干潟で採集されたヨコヤアナジャコ。額角(rostrum)の形態と頭胸部背面のひょうたん模様から,ヨコヤアナジャコとみて間違いないが,yokoyai かどうかはクレード B, C, D を比較してみないと判断できない。また,ヨコヤ氏の描いたスケッチ(図 11)は,このアナジャコが yokoyai かどうかを判断する基準(standard)としては使えない。では専門家に相談すればよいではないかという人は多いだろうが,平野さんを通じて自称専門家の判断がいかに不正確かを痛感させられた。私は,客観的な証拠(データ)に基づいて自分たちで判断する道(普通の自然科学的方法)を選んだ。

図 17.名蔵川の泥干潟で採集されたスナモグリ。種名は不明だが,いくら接写であっても,写真だけから図 15 に示した個体と同種と断定することは難しい。頭胸部背面の溝は,幅の広い小舟のような感じである。ひょうたん模様ではない。

図 18.名蔵川の泥干潟で採集されたスナモグリ。頭胸部背面の模様(溝)はひょうたん型ではなく,幅の広い小舟型。額角(rostrum)は大きくて鋭い。外見でも見分けやすいので,和名としてヨツバキイロスナモグリ(仮称)と名付けた。

図 19.名蔵川の泥干潟で採集されたヨコヤアナジャコ(♂)。頭胸部背面にはひょうたん溝があるのでヨコヤアナジャコには間違いないが,図 10 や図16 に示した個体とは,頭胸部背面のひょうたん模様に明らかな違いがある。つまり,図 19 の個体では,ひょうたんのくびれが図 10 や図 16 の個体に比べて強い。また眼柄(eyestalk)外側にある「襟(えり)」(起源はアナエビの第 2 触角の基部にある大きな棘ではなく,頭部前面にある側方額角(lateral rostrum)だろう)との間隔が広いために,上から眼柄の先端がよく見えるのが特徴である。ひょうたんのくびれから上の部分も図 10 や図 16 の個体に比べて,相対的に長い。この写真だけで新種とは認定はできないが,少なくとも図 10 や図 16 の個体とは形態的に違う個体が混じっていると言えるだろう。眼柄の脇にある棘(spine)も 3 つある個体,2 つある個体,ひとつしかない個体が混じっている。雌雄差のせいではないだろう。

図 20.宮良川のマングローブ。2024 年 6 月 20 日(木)は,石垣島の宮良川にあるマングローブに行った。2024 年 6 月 20 日は,石垣島では干潮(low tide)は 12 時 28 分。潮位は 32 ㎝。大潮(spring tide)が近い。11 時ごろの撮影ではないかと思う。まだ潮はあまり引いていない。

図 21.泥干潟に現れたハクセンシオマネキ(宮良川)。ハクセンシマネキは干潟の砂地に住んでいる。潮が引くといっぱい巣穴から出て,摂食やオス同士のけんかをしている。シオマネキは,生息地によって抱卵する時期が多少異なるかもしれない。この辺り(砂地)はまだ歩きやすい。

図 22.泥干潟に現れたオスのヤエヤマシオマネキ(宮良川)。底質(substrate)が泥に変わると,住んでいるシオマネキの種類も変わる。泥(silt やclay)が堆積した干潟には,ヤエヤマシマネキやヒメシオマネキが住んでいる。長靴を履いて歩くと足を取られる。

図 23.軟泥層(ヘドロ)の上に現れたヤエヤマシオマネキのオス。近くにはミナミトビハゼやオサガニもいる。泥の表面は酸素があるので茶色になるが, 2~3cm も掘れば灰色の RPD zone になる。RPD zone や anoxic zone でなぜ動物が生きられるかというと,動物が歩脚(pereiopod)を使って泥に穴をあけて自分が動ける空間を作るからである。一時(今も?)泥に住むアナジャコには水質の浄化作用があると主張する人たちがいたが,アナジャコが多く住んでいる干潟でも,巣穴の中で好気性のバクテリアが爆発的に増加するという話は聞かない。富栄養化が抑制されるという証拠もない。生態学者は環境問題について,根拠の希薄な証拠を持ち出して話を作るものだから,出された仮説は全然説得力がない。確たる根拠のある証拠(たとえば好気性細菌によるに有機物の分解)に基づいていたとしても,「水増し仮説」を立てる。環境汚染が生物に影響を与えていると言っても,汚染前の姿(コントロール,ある意味正常な姿)がないので,本人が勝手に思い込んでいるだけと疑いたくなる仮説も多い。日本化学工業協会から私に委託された研究テーマ(5~6 年続いたプロジェクト)は,生物の正常な姿(どこでどんな生活をしているか)を明らかにしてほしいということだった。

図 24.宮良川(石垣島)河口の泥干潟。20 ㎝から 30 ㎝の厚さの軟泥層(ヘドロ)が堆積し,軟泥層の下にはサンゴ礫が現れる。アナジャコの巣穴はたくさんあるが,何度泥を掘ってもアナジャコやスナモグリは採集できなかった。よく似た泥環境が南西諸島の多くのマングローブ域に広がっている。何度挑戦してもアナジャコやスナモグリを採集できないので,最後の方は遠くから干潟を見ているだけになってしまった。

図 25.河口に堆積した軟泥層(宮良川)。軟泥層は 20 ㎝も堆積すれば,長靴と泥の間に空気の層がなくなるので,すごい力で引っ張らないと,長靴を泥から抜くのが難しくなる。足の方が先に抜けてしまうので,泥に残った長靴に足を入れ直すところでバランスを崩して,足はヘドロの中に‘ぼちゃん’と埋まった。結局下半身は泥だらけになった。宮良川の干潟ではさすがに採集が嫌になって,近くのコンビニにある水道で足と衣服を洗ってからレンタカーを返し,離島桟橋行のバスに乗った。次は西表島である。私はこんなことが楽しくて生きている。