
1.Introduction
生物多様性(biodiversity)や進化生物学(evolutionary biology)の研究では,地球上の様々な環境に生息する動物や植物が対象になる。現在進行中の研究プロジェクトのテーマは,十脚甲殻類の進化と高次機能の発現機構である。研究を進めるためには,研究対象とする十脚甲殻類の属(genus)や種(species)の名前を知らなければならない。生物名は,属の上位の分類群である「科」(family)や「下目」(infraorder)等にもつけられている。
日本では,生物の名前に表記は,一般には和名(Japanese name)が使われている。「ヒト」とか「モンシロチョウ」とか「アカテガニ」は,和名(固有名詞)である。和名をつけるための規約はないが,カタカナ表記が原則になっている。中学や高校の生物の教科書に出てくる生物名は,「ヒト」を含めてすべてカタカナで表記されている。生物の教科書に,「ひと」,「もんしろちょう」,「あかてがに」と書かれると,時代は一気に明治にさかのぼる。「赤手蟹」と漢字で書くと,さらに江戸時代にさかのぼる。もし高校の教科書に「蜚蠊」とか書かれていたら,生物の教科書は漢文の教科書に変身するかもしれない。ちなみに「蜚蠊」は「ごきぶり」と読む。
生物名には,日本語(主にはカタカナ)で表記する和名の他に,論文等で使用する学名(scientific name)がある。学名は,リンネの2命名法に基づいて表記するという約束がある。例えば,モンシロチョウはPieris rapae,アカテガニはSesarma haematocheirという具合である。PierisやSesarmaは属(genus)の名称,rapaeやhaematocheirは種(species)の名称である。リンネのころは,学名はラテン語を使うという約束があったが,今は自由になっている(ラテン語を理解できる人は皆無ということもある)。例えば,ベンケイガニの学名は,Sesarma intermediumでもS. intermediaでもS. intermediusでもよい。属名と種名は固有名詞と混同を避けるために,両者はイタリックスで表記される。専門家もアマチュアも,語尾に「i」とか「ensis」をつければラテン語になると思っているらしく,okashina-ensisとかinchiki-i とかkimoiみたいな種名が飛び交っている。百人一首,カルタ取り,観葉植物の世界で生活する人たちならいいが,そんな種名を使って生物学の研究を進めるというのは,さすがに時代錯誤も甚だしい。
生物の命名に関して大きな問題は,種名の表記に固有名詞(人名)が頻繁に使われることである。例えばヨコヤアナジャコはUpogebia yokotai,サカイツノガイヤドカリはPylocheles sakaii,オオバヤシトゲヒゲトラカミキリはDemonax ohbayashiiとなっている。人名ではないが,Inachis geisha(クジャクチョウ)なんていうのまである。どういう基準や経緯で人名が入るか不明であるが,人名を入れた学名は,十脚甲殻類や昆虫の世界では特に多く見受けられる。しかもsakaiとかohbayashiとか,特定の人物の名前が多くなっているのが特徴である。
近代的な博物学(natural history)が始まったのは,リンネ(Carl von Linné;1707–1778)の時代である。そのころは,種を記載することが生物学の主要な目標だった。そして採集された生物には名前を付けなければならない。生息場所とか,種の特徴を考慮して学名をつけた分類学者もいただろうが,実際には権威者や自分自身に献名し,ご機嫌を取る者も多かったのではないか?
種名や属名に固有名詞である人名を付けるという行為は,一般の人たちにとっては大きな関心事にはならない。しかし,分類学者の中には生物に名前を付けることに生きがいを感じている者もいる。かつて東京大学植物学教室で,トガクシソウの命名をめぐって,伊藤篤太郎が矢田部良吉(1851-1899)教授に破門された事件(破門草事件)があった。もともとは伊藤が記載した新種を,ロシアの植物学者マキシモヴィッチがYatabea japonica Maxim. として献名しようとした。伊藤はこの動きをいち早く察し,Ranzania japonica (T. Itô ex Maxim.) T. Itô(1888)としてイギリスの植物学のジャーナルに先に発表した。矢田部良吉はこれに激怒し,伊藤篤太郎を植物学教室に出入り禁止にしてしまった。マキシモヴィッチは, RanzaniaとかTogakushia(これは私の造語)という属(genus)にしておけばよかったのだろうが,矢田部良吉に忖度してYatabeaなどと固有名詞(人名)をつけたものだから,伊藤篤太郎が怒るのも当然だったと思う。命名者の名前は種名の後に表記される(例えばT. Itô ex Maxim.)。個人の名誉欲は,それで十分満足されるのではないかと思う。矢田部良吉の欲の深さは,度を越えているように思えるのだが・・・。
大学に入学して,2年次(初年度だったかもしれない)に系統分類学の授業があった。授業の内容は.かつての東京帝国大学動物学教室の講義とほぼ同じだったと思う。進化の観念が抜け落ちているので,それぞれの動物群(門や綱)の基本的な体の仕組みを解説していた。恐ろしく退屈な授業だったが,この講義(関口晃一先生)を聴講してひとつだけ記憶に残ったことがある。それは「生物種の記載に人名を入れる例があるが,ああいうのは良くない。」と言われていたことである。関口先生の頭には,ひょっとして丘浅次郎(東京高等師範学校)がホヤの新属・新種に自分の名前(浅次郎)をつけて「Asajirella gelatinosa Oka, 1891」と命名したことが頭にあったかと想像したが,さすがにこれは私の邪推だろう。
伊藤篤太郎(いとうとくたろう)の例が示すように,献名と称して種名や属名に権威者の名前を入れると,採集や分類の過程に関わってきた者との間にトラブルが起きやすい。種や属を同定したのは自分だから,種名や属名に自分の名前を付ける権利がある,などという約束ごともない。
私が一番言いたいことは,自分の競争相手の人物の名を付けられてしまったら,それ以後の研究はものすごくやりにくくなるということである。伊藤篤太郎がトガクシソウを一所懸命研究しても,Yatabeaと名前がついていることで,成果は矢田部良吉に献上する感じになる。矢田部からすれば,「いやいや伊藤君,私の名前が付いたトガクシソウを研究してくれて,大変有難う。」という勝ち誇った気持ちになるかもしれない。そして「あの植物はな・・・,実は私が命名したんだぜ!」という高揚した気分も生まれるだろう。こんなことを陰日向で聞かされたら,伊藤篤太郎はどう思うだろうか?私には,伊藤篤太郎が矢田部良吉に対してどんな感情を抱くか,痛いほどよくわかる。
周囲から見ていると,そんな些細なことで喧嘩するより穏便に済ませたら,と思うかもしれない。権威者に仲裁を依頼することは,多くの人たちが実行している。しかし,安易に権威者・権力者を頼ることは危険である。権威者は,双方の主張の真ん中で線引きをする。よく考えてみていただきたい。権威者とは多くの人がそう思うから権威者になれるのである。片方の主張にだけ味方したら,もう一方の主張から激しい反発を食らい,権威者の地位には就けない。つまり八方美人でないと権威者にはなれない。だからこそ,権威者の持つ倫理観には恐ろしい判断が潜んでいる。
権威者による線引きは,実に理不尽な根拠の上に立つ。そんな判断を承諾するのは負けたのと同じことである。あとはうじうじと生きる人生が待っている。そんなみじめな結果になるならば,権威者など頼らず,自分の持てる力をすべて出し切って争う方がよい。
多くの大学の理学部では,最近は系統分類学の研究室はほぼ消滅しているので,種や属の命名をめぐって大きな問題は起きていないようである。唯一私の研究室(最終的には進化生物学研究室)では,アナジャコ類の分類と名称に関して大きなトラブルが起きた。トラブルを起こした張本人は,・・・他ならぬ私自身である。この事件はまだ終息していない。近いうちに決着をつけなければならない。自分で言うのも何だが,私は伊藤篤太郎よりもはるかに始末が悪いと思う。あっちこっちで「破門」を食らっている。しかし,破門を食らえば,ますます自分のやりたいことを思い切りできるようになる。
最後に,私はアマチュアや観葉植物の世界で,学名に人名をつけてはいけないなどと言っていない。学名に人名をつけて学問になる世界,例えば,分類学会,甲殻類学会,観葉植物の学会では,自由にやっていただければよい。ただ,私は百人一首やカルタ取りの世界を自分の学問の中に持ち込みたくないだけである。一方では,生物の分類を必要とする分類学者,農林水産関係の研究者や技術者とは接点を維持して行きたい。それらの方々には,自然に生きる多くの生物の姿(人間も含めよう)を紹介することで接点を保ち続けたい。視点が違うだけで喧嘩別れになるのは,お互いにメリットがない。
2.記事の執筆と編集
三枝誠行(生物多様性研究・教育プロジェクト,常任理事)
増成伸文(生物多様性研究・教育プロジェクト,常任理事)
3.参考文献
・Forest, J. (1987) Les Pylochelidae ou ’Pagures symétriques’ (Crustacea Coenobitoidea). Mém. Mus. nat. D’Hist. nat., Paris. 3 (Zool.): 1‒254 with 9 plates.
・Futyuma, D.J. 1998. Evolutionary Biology. Third edition. Sinauer Association. Massachusetts.
・Glaessner, M.F. (1969) Decapoda. In: R.C. Moore (ed.) Treatise on Invertebrate Paleontology. Part R, Arthropoda 4, Vol. 2, pp. R399‒R628. University of Kansas, Lawrence, Kansas, and Geological Society of America, Boulder, Colorado.
・Herrick, F.H. (1895) The American lobster. A study of its habits and development. Bull. US Fish Comm. 1895: 1‒252 with 54 plates.
池田嘉平・稲葉明彦 1971. 日本動物解剖図説。森北出版。
・McLaughlin, P.A., and R. Lemaitre (2009) A new classification for the Pylochelidae (Decapoda: Anomura: Paguroidea) and description of new taxa. Raff. Bull. Zool. Suppl. 20: 159‒231.
・三宅貞祥 1982.原色日本大型甲殻類図鑑(Ⅰ)保育社。
・Richter, S., and G. Scholtz (1994) Morphological evidence for a hermit-crab ancestry of lithodids. Zool Anz 233: 187–210.
・椎野季雄 1964. 動物系統分類学 7(上) 節足動物(Ⅰ)総説・甲殻類。中山書店。
・内田亨 1965. 動物系統分類の基礎。北隆館。
・Williams, A.B. (1995) Taxonomy and evolution. In: J.R. Factor (ed.) Biology of the Lobster: Homarus americanus. Academic Press, San Diego.
・山田真弓・西田誠・丸山工作 1981. 進化系統学。裳華房。

図 1.土佐湾の深海の底引網で採集された魚介類。この時(2020 年 11 月だったか)は,水深 250m あたりの底引きを行ったと思う。底引網で獲れた魚介類はトロ箱ひとつあたり 1 万 5 千円で購入した。お金を出さないと,網を底まで下ろさずに引いてしまうので,魚(メヒカリやレンコダイ)が入る。トロ箱の中の魚類や甲殻類は,強烈な臭い(腐敗臭とはちょっと違う)を発する。エビなどは冷蔵して持ち帰っても,食べようという気持ちにならない。
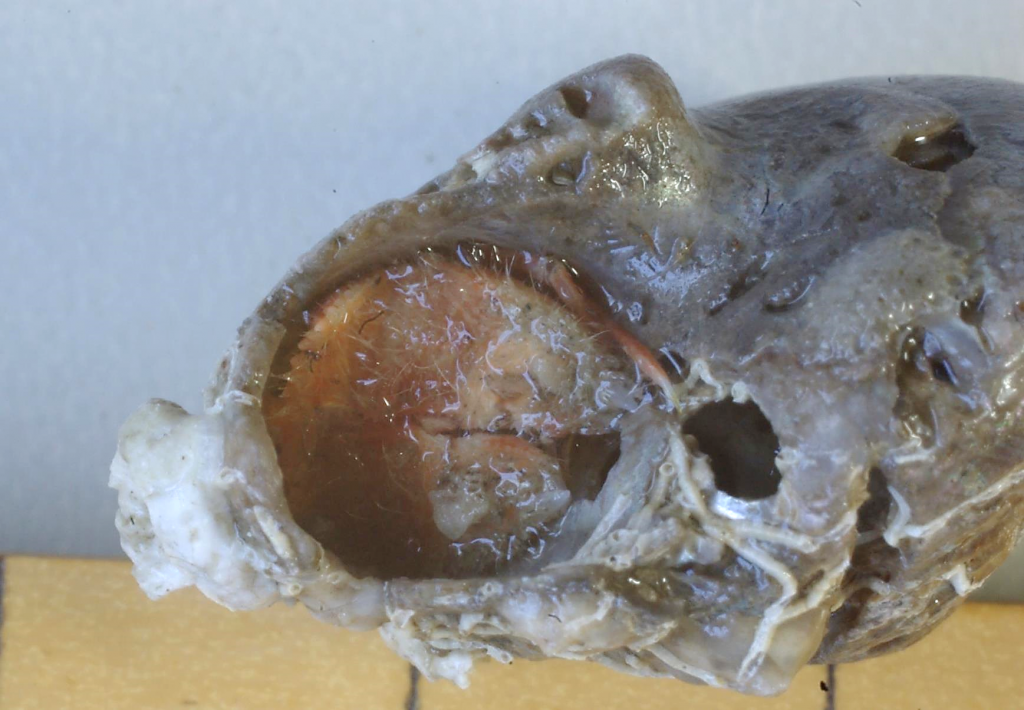
図 2.壊れた貝殻の中に入っていたカルイシヤドカリ(Pylocheles mortensenii)。両方の鋏脚(propodus と dactylus)をそろえて円形の穴をふさいでいる。このような穴のふさぎ方から,カルイシヤドカリの鋏(chela)は胸部に向かって下側を向くことになった。カニや他のヤドカリは横方向を向いている。
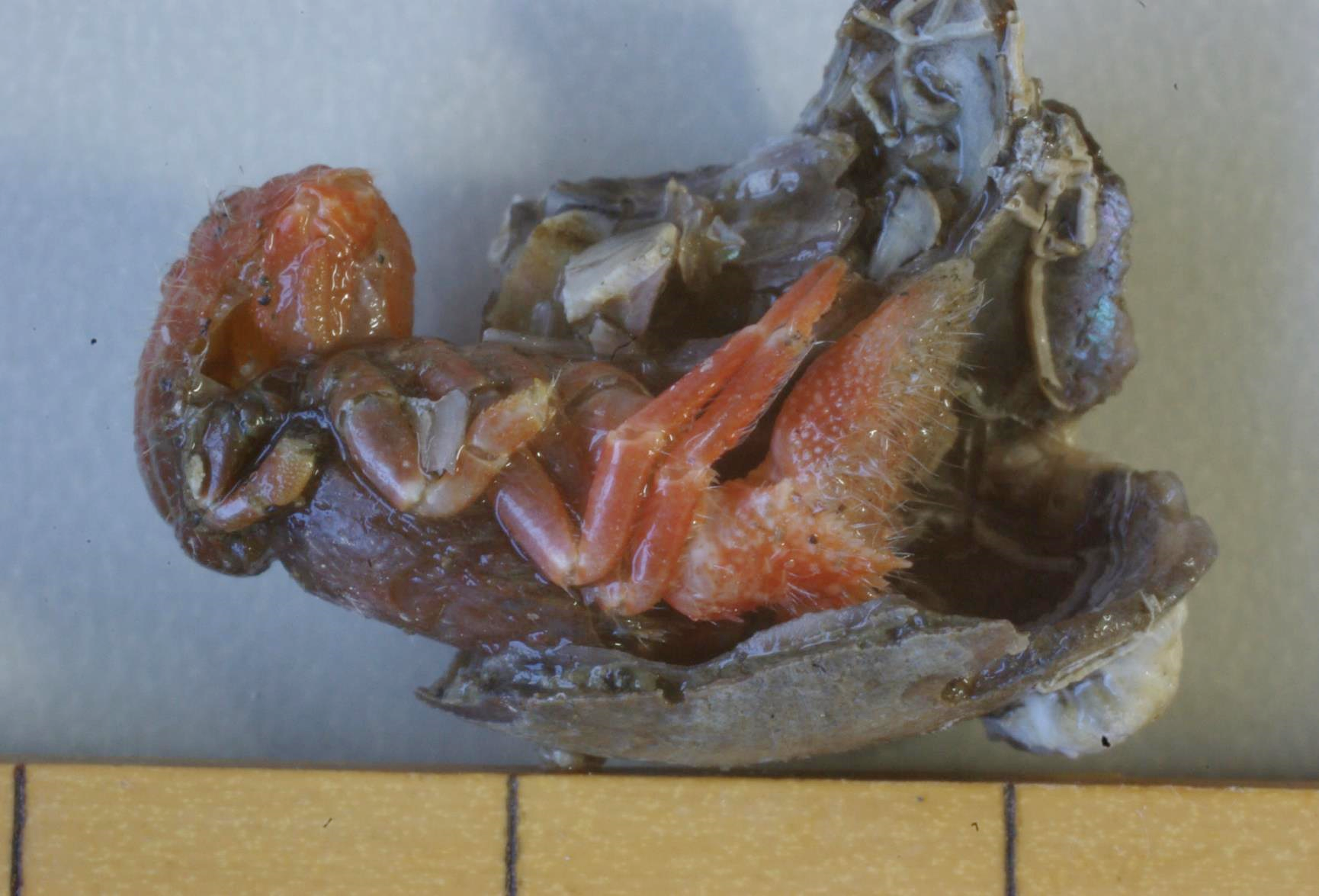
図 3.壊れた貝殻の中に入るカルイシヤドカリ(Pylocheles mortensenii)。自然に壊れた貝殻をさらに壊して全身が見えるようにした。体長は 3 cm ほど。ヤドカリの中では非常に古い時代(ジュラ紀と白亜紀の間あたり)に出現した。土佐湾には最も古いヤドカリ(Cancellocheles)も生息している。
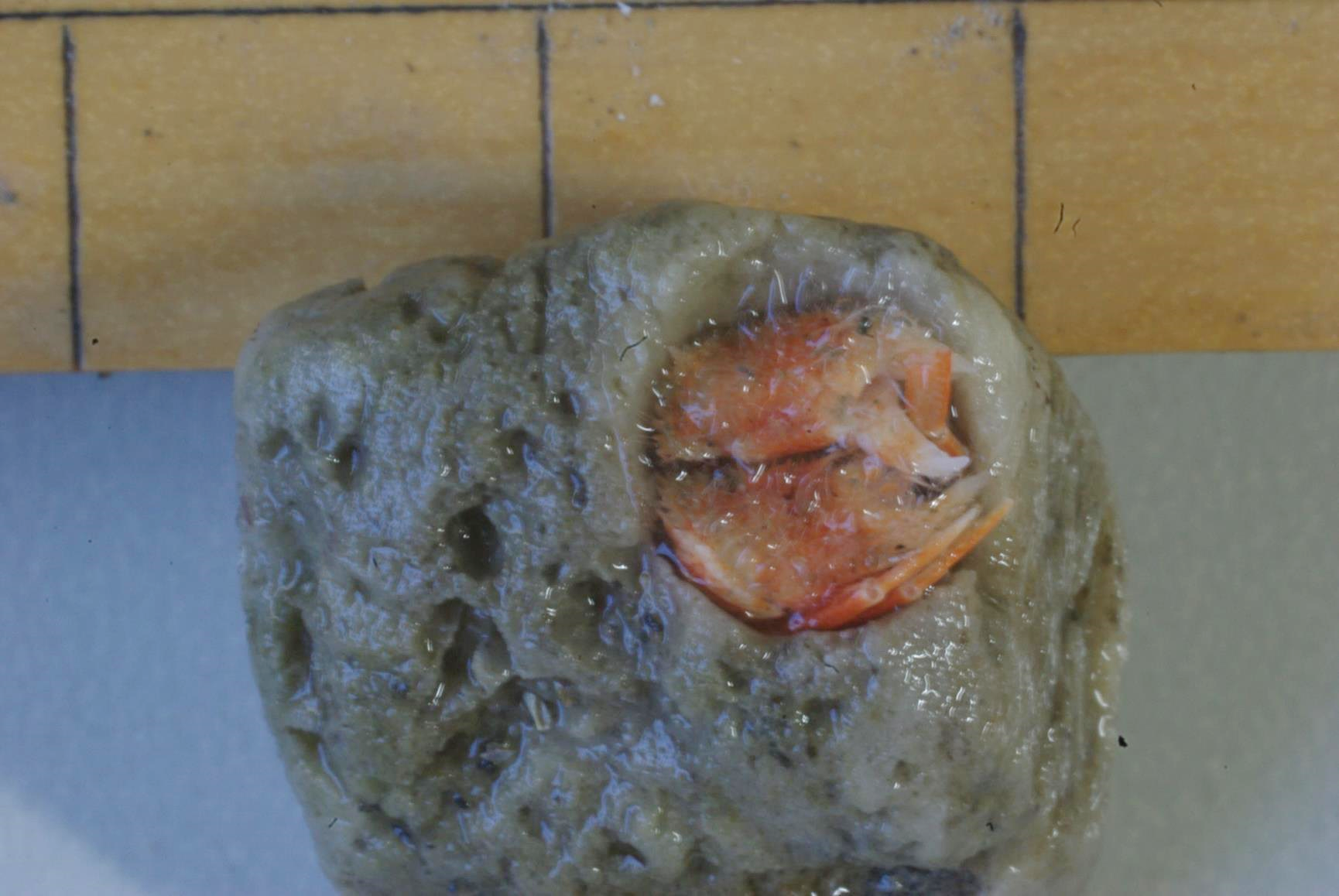
図 4.クロシオカルイシヤドカリ。軽石(直径 2 cm)に穴を掘り,両方の鋏脚を使って穴をふさいでいる。すでに他の人が行った遺伝子解析と照合した結果,学名は Pylocheles mortensenii で間違いはない。和名をつけるとすれば「クロシオカルイシヤドカリ」ではどうか?いずれにせよ,今は遺伝子のデータがないと分類に関する議論は平行線をたどる。しばらくは,互いに様子見の関係(意味不明だが・・・)で研究を進めるのがよいだろう。

図 5.軽石の穴に入るクロシオカルイシヤドカリ。穴の入り口は鋏脚の先を下に折り曲げて全体をふさげるようになっている。自分で掘ることもできるだろうし,他の個体が使った巣穴を利用して穴を拡充するということもあるだろう。第 4 歩脚と第 5 歩脚の先端には滑り止め(rasp)がある。

図 6.コゲチャカルイシヤドカリ(Pylocheles atrosanguineus)。atro+sanguineus は,ラテン語で「暗い赤」の意味。私のノートが間違っていなければ,Pul-04(Kal-04 の間違い)は, カルイシヤドカリ(Pylocheles)の 1 種であるが,mortensenii ではない。Pylocheles は,形態上の個体変異(intraspecific variation)が大きく,形態では種の判定が難しい。分類学者は強引に分けてしまうか,紛らわしい個体は初めから分類の対象からはずしている。(つまり形態的に違いのある個体のみを「種」として認めているのであって,遺伝子解析の結果から 2 種に分かれると言っても,見向きもしない。だから,分類学者の言うことを鵜呑みにしていては,系統学的・進化学的研究に大きな支障をきたすことがある。特に,中生代の白亜紀から新生代にかけて地球上に
出現した生物の系統関係(phylogenetic relationship)については,形態学的な研究よりも分子系統学的研究の方がずっと信頼できる。Pul-04 は,Forest
(1987)が記載した Pylocheles のどれかと思われるが,古い文献には遺伝子解析のデータがないので照合できない。分類学者に任せたら,呪い師の世界に逆
戻りだ。さらに Yatabea のように,(伊藤篤太郎からすれば)とんでもない奴の名前が候補になれば,自分の研究は邪魔されていると思うだろう。そんな
屈辱的な状況になる前に,さっさと自分で新しい名前をつければよいのである。また,Pylocheles sp. にすると古い分類学者との関係が断ち切れない。

図 7.クロシオカルイシヤドカリ(Pylocheles mortensenii)。カルイシヤドカリは,2019 年から 2021 年の 3 年間で 100 匹以上採集することができた。水深
350m から,浅いところでは 30m ぐらいの海底から採集された。太平洋の海底には広く分布しているのだろう。
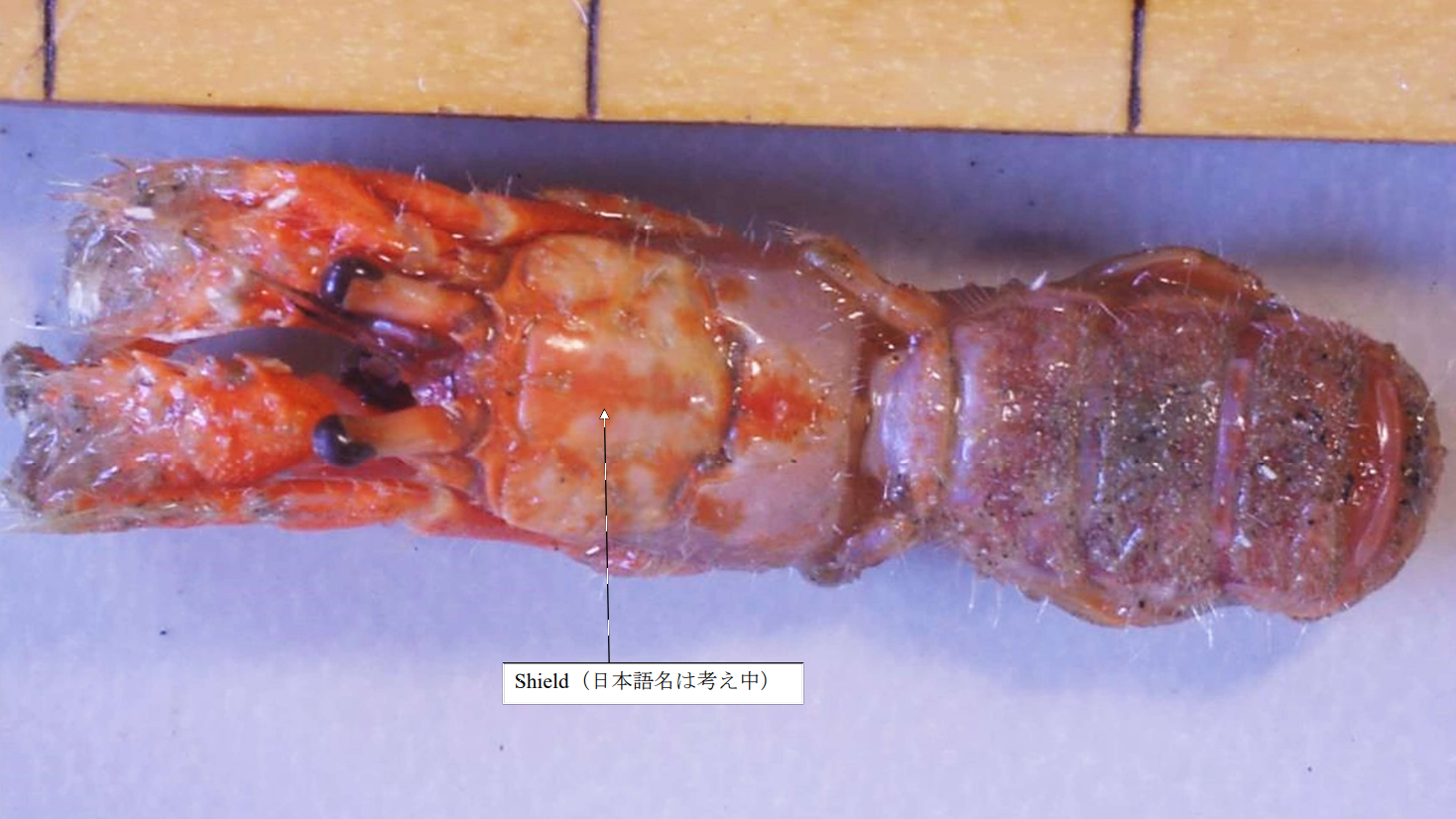
図 8.トガリツノガイヤドカリ(Trizocheles scaphopodae)。ツノガイ(Scaphopoda)の殻に入る Trizocheles 属のヤドカリだが,数は少ない。いままでに 10匹程度採集されている。ツノガイヤドカリは土佐湾では 2 種類記録されているが,遺伝子解析では今のところ 1 種類しかいない。和名はツノガイヤドカリ。分類学者は甲羅の形態に注目して種を決めているが,胸部背面は腹部と同様に脱石灰化が進んでいる。実際に得られた標本は個体変異が大きく,胸部背面の形態や眼柄の太さ・長さでは種の区別は難しい。しかもアルコール漬け標本は見分けにくい。トガリツノガイヤドカリは 土佐湾から複数の種類が記録されているが,1 種類ではなかろうか?遺伝子解析の結果が形態的観察と一致しない場合には,遺伝子解析に基づく新しい学名をつけるよ,と公言した方がよい。Trizocheles scaphopodae なんていう学名はどうだろうか?人によりけりではあるが,話し合いで合意に至る可能性は少ない。

図 9.トガリツノガイヤドカリ(Trizocheles scaphopodae)。
遺伝子解析では,図 9 に示したツノガイヤドカリは,図 8 に示したツノガイヤドカリと同じ種になるが,外皮(exoskeleton)の色は大きく異なる。眼柄(eyestalk)の形態も違っており,外部形態の観察では両者は別種にされる可能性が高い。Forest (1987)(フランス語で書かれている)には世界中から集められたカルイシヤドカリとツノガイヤドカリのスケッチが掲載されている。Forest (1987)の論文では,ツノガイヤドカリはアルコールかホルマリン固定標本に基づき,胸部背面の shield の形態,額角の有無,眼柄の太さや長さなどから種を分類している。
私がカルイシヤドカリやツノガイヤドカリをよく調べた結果では,両種とも
胸部背側にある shield は脱石灰化(decalfication)が進んでおり(特に側面は顕著)外観では種の判別は難しい。分類学者は,液浸標本ですら分類できると主張するが,私はそんな議論に加わる意思は持ち合わせていない。私は,系統関係を量的・客観的に推定できる分子系統解析も参考にして,種を決めている。古典的方法と分子系統解析の結果は合致しないことが多い。もし完全に合致したとすれば,それは分類学者が,塩基配列をシークエンスする前に,形態では分類できない個体をあらかじめ除いてしまったからである。
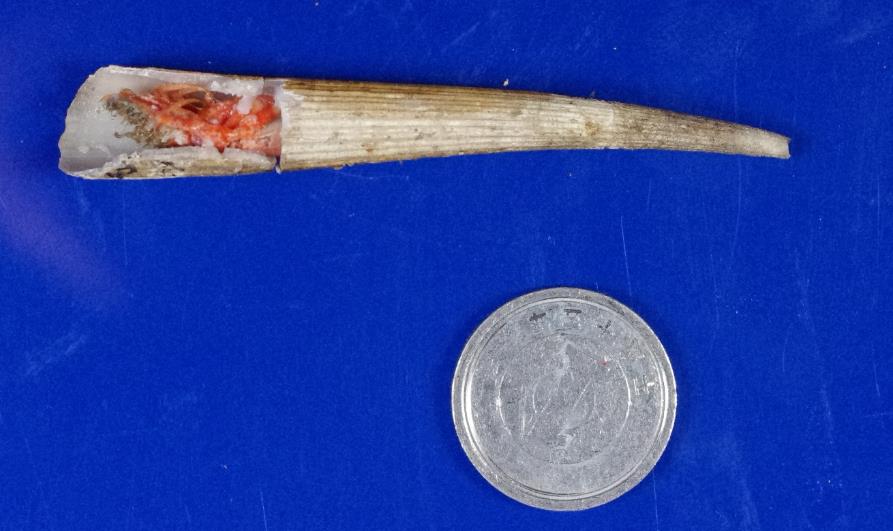
図 10.ツノガイに入るツノガイヤドカリ。1 円玉はスケール(直径 2cm)。

図 11.アメリカン・ロブスター(和名)。学名は Homarus americanus。日本の沿岸には生息してない。多くの十脚甲殻類の起源を探してゆくと,多くの種類(抱卵亜目)は,ロブスターの祖先から進化したことがわかる。筋肉は,鋏(propodus)と腹部(abdomen)にたくさん入っていて,味もすごく良い。
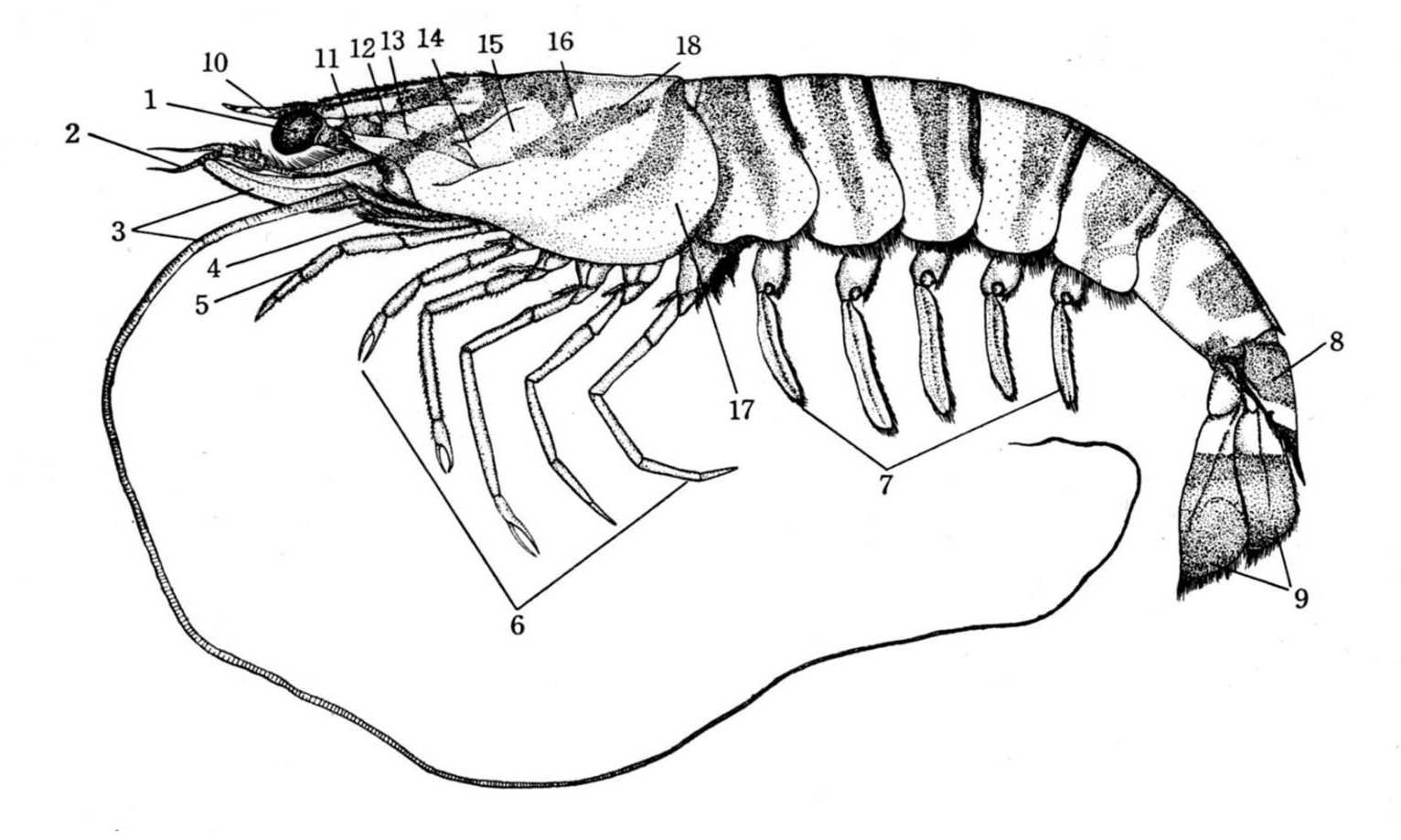
図 12.クルマエビの全形(池田・稲葉「日本動物解剖図説・クルマエビ」から転載。クルマエビヤウシエビは,ロブスターとは違った系統である。十脚甲殻類は,二畳紀の終わりから三畳紀の始めにかけて起きた海産動物の大量絶滅の後に,絶滅を逃れたアミ類が海底に下り,底生生活(benthic life)を送る過程で進化したのだろう。もちろん異論はある。体もそのころ(三畳紀)に大型化したと思われる。クルマエビ(根鰓亜目)がなぜ原始的な種類と言えるかというと,ロブスターは受精卵を腹部に抱えて幼生が孵化するまで保持するのに対し,クルマエビは(おそらく満潮時に)砂泥底から泳ぎ出て,卵を海中に勢いよくばらまくからである。受精は卵がばらまかれた直後に起きる。形態や行動の研究を長く続けていると,受精卵を腹部に付着させる種類(抱卵亜目)は,三畳紀(Triassic)の初期に,未受精卵を海中にばらまく種類(根鰓亜目)から進化した可能性が高いと思えるようになる。では,ロブスターも含めて,原始的な十脚甲殻類はすぐに絶滅するかというと,そういうことはない。進化した生物でも早い段階で絶滅することもある。
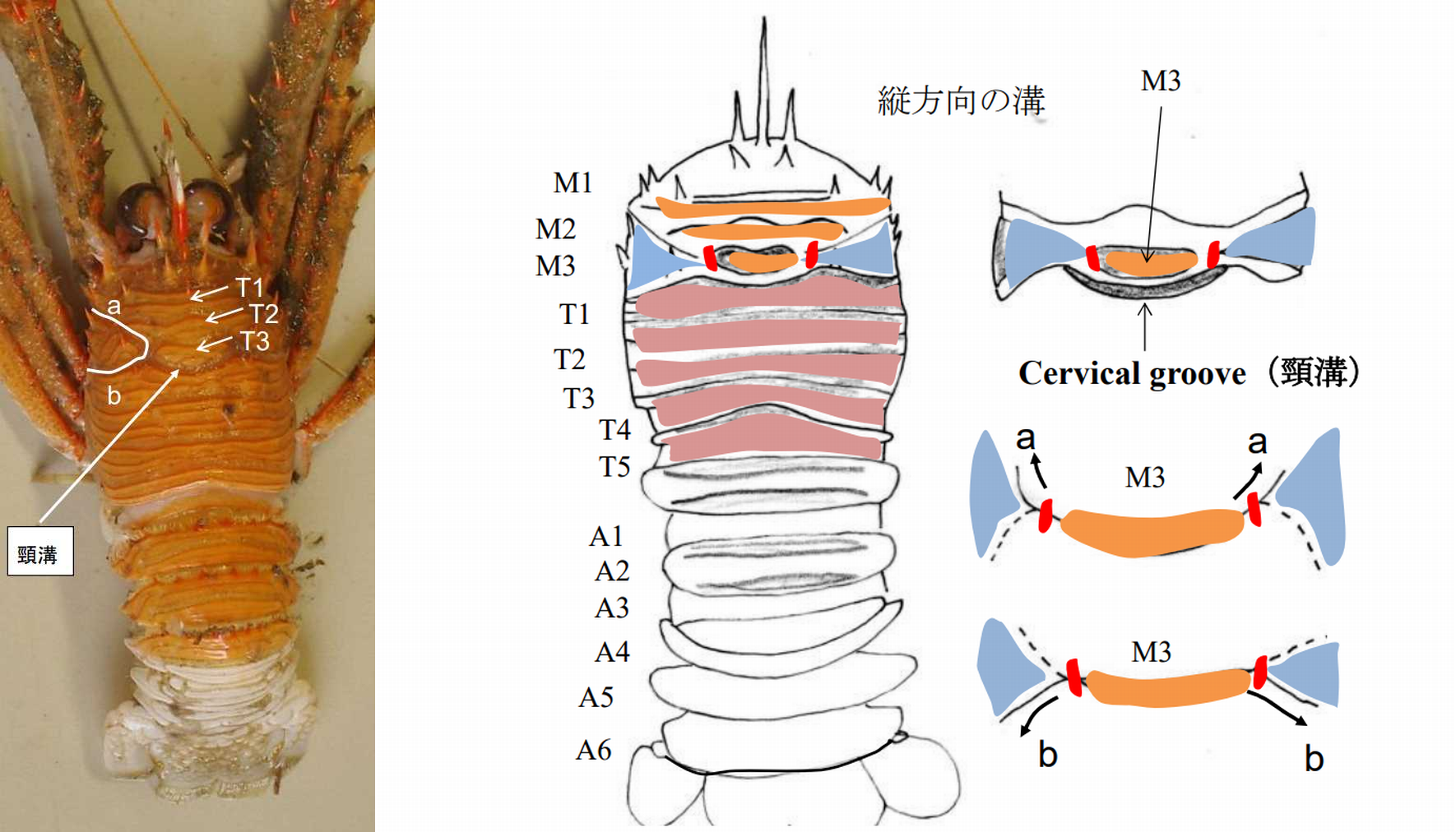
図 13.オオコシオリエビに残る十脚甲殻類の体節(segment)。
根鰓亜目,抱卵亜目ともに十脚甲殻類の体節(segment)は,腹部(A1–A6)を除いて不明瞭になっている。ところが,オオコシオリエビでは体節が古い形のまま残っている。M1 から M3 は,第 1 から第 3 顎脚に対応するプレートであり,T1 から T6 は胸部付属肢に対応する(T1 のプレートは鋏脚に対応)。十脚類は同じ甲殻類のアミ目から進化したと言われているが,アミ目はすでに十脚類の根鰓亜目とよく似たプレート構成になっている。つまり,十脚類の祖先はアミ目ではなく,もっと別のグループだったかもしれない。コシオリエビのプレート構成を見ると,ワラジムシとかスナホリムシ(ともに等脚類)が近いように思える。Cervical groove の有無も面白い。もう少し証拠が欲しい。
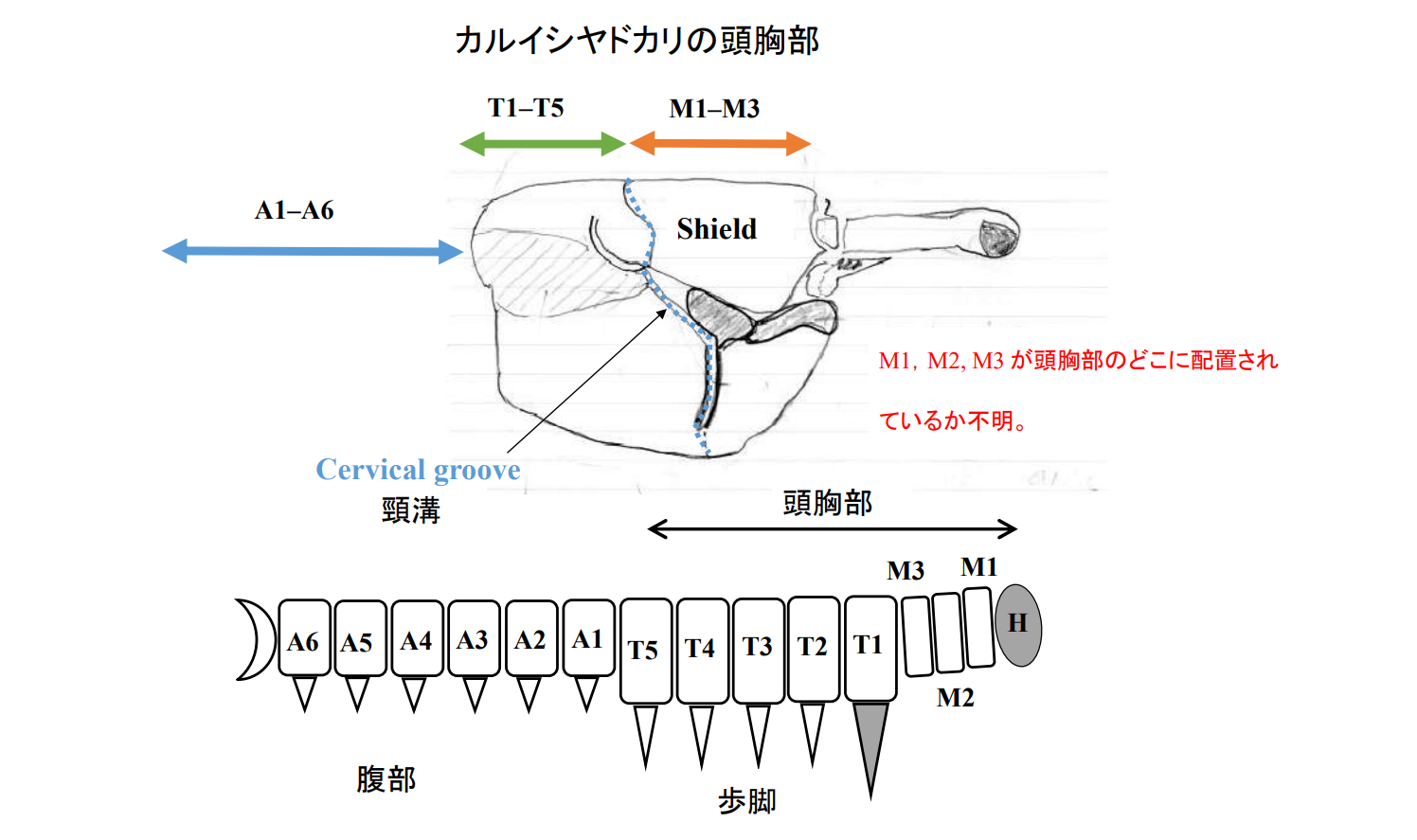
図 14.十脚甲殻類の体の外部形態。十脚甲殻類は,同じ節足動物門(Phylum Arthropoda)でも,昆虫とは大きく異なっているところがある。ひとつは,昆虫類は付属肢が 3 対(計 6 本)に対し,十脚甲殻類は 5 対(計 10 本)ある。なぜ昆虫の肢が 3 対で,十脚甲殻類は 5 対かという質問に答えるのは難しい。ともに最初は,たくさんの肢があったが,進化の過程で昆虫は 3 対の肢をもったグループが,十脚甲殻類は 5 対の肢をもったグループが生き残ったのだろう。もう一つの大きな違いは,昆虫では顎脚(maxilliped)に対応するプレートが欠損しているが,十脚甲殻類では顎脚プレート(M1–M3)が残っている。十脚甲殻類では顎脚を作る遺伝子が発現しているだろうが,対応するプレートは不明瞭である。

図 15 .テナガエビの外部形態。西表島の古見岳に登る登山道沿いの川で採集した。抱卵メス。抱卵亜目のエビ(prawn)の中でこれだけ大きい種類は珍しい。内湾ではクルマエビヤウシエビのような根鰓亜目(こんさいあもく)のエビが残り,抱卵亜目の方はみな絶滅した可能性がある。

図 16.テナガエビのオス。古見岳に行く小道沿いの川(川の名前は知らない)で撮影。テナガエビのオスはとんでもなく長い鋏脚(cheliped)を持っている。鋏脚は,指節(dactylus)と掌節(propodus)が向かい合ってハサミになっている。十脚類の鋏脚は,指節が掌節の上側にあるが,テナガエビはdactylus が propodus の下側についている。鋏脚(胸部第 1 歩脚)のどこかの節が成長に伴ってねじれた結果,先端の節が反転したのだろう。

図 17.テナガエビのオス。古見岳に行く小道沿いの川で採集した。クルマエビヤウシエビのような根鰓亜目のエビや,テナガエビのような抱卵亜目のエビは甲羅の上に cervical groove(頸溝)が現れない。また,頭胸部の先端にある縦方向に伸びる茶褐色の長い突起は,ロブスターやアカザエビの頭部先端に見られる凶悪な突起に似ている。この突起(額角)はクルマエビにもあったと思う。西表島にはテナガエビは 2 種類いるかもしれない。

図 16.古見だけに至る登山道の入り口付近。古見と美原の中間に,古見岳登山道がある。県道 215 号の脇に車を止めて,小川を何度も横切りながら(橋はない)渓流沿いを歩く。215 号の入り口から数キロメートルは平たんな道が続いていて,水生生物の観察には適している。しかし,水生動物を採集する予定が,昆虫採集の装備になってしまった。手前に小川があり,石がごろごろしている。石の隙間をよく見ると,テナガエビの長い鋏脚が見える。

図 17.登山道での採集風景。川を何度も横切るので,長靴は必需品。この道では昼も夜もサキシマハブに出くわしたことはない。どこに行ってもそうだが,生物採集となるとどうしても昆虫の方に目が行く。しかし,この場所(215 号から 500m ほど入ったところ)は少し開けていてチョウがいそうなものだが,不思議と少ない。イシガケチョウ,クロセセリ,ウラギンシジミ,ルリウラナミシジミ,ヒメウラナミシジミ程度しか観察できない。

図 18.川沿いで休憩中。県道から 1km ぐらい入ったところ。まだ平坦な小道が続いている。サワガニを採集したところと思われる。西表島には数種類のサワガニがいるが,石垣島の山の中にいるでっかいサワガニ(種類は知らない)はいない。右側の小川の少し深いところにテナガエビがいる。

図 19.テナガエビのオス。右の鋏脚が取れてどこかに行ってしまった。オスの鋏脚は何でこんなに長いのか,理由はよくわからない。遠くにあるエサを取るためならオスメスともに鋏脚が長くなって良いと思うが,メスはオスのように長くない。すぐ自切するような鋏脚はケンカにも役立たない。

図 20.十脚甲殻類のメスの腹部にある付属肢。十脚甲殻類の腹部にある付属肢(腹肢)は遊泳肢とも呼ばれる。メスの腹肢は担卵肢となっていて,外肢と内肢の両方,もしくは片方に生えた担卵毛に受精卵が付着する。C で示した写真には担卵毛に受精卵が付着したところが写っている。十脚類の抱卵の詳細については,おいおい述べて行きたい。

図 21.西表島の自然林内の風景。川にはサワガニ(多分 1~2 種類)とカノコガイ(だったっけ?)がたくさんいる。また,もう少し下流には,ユゴイ(淡水魚)がたくさん集まっているところがある。夜になると,テナガエビの他に,オオウナギが出てくる。オオウナギは追いかけると,水から出て砂利や小石の上を逃げ回る。ウナギはまず見ない。夜に行くならば,川を歩いて登らないと 5 分も経たないうちに小道を見失い,パニックに陥る。

図 22.西表島の自然林。手前の大きな木はオキナワウラジロガシだろう。島の西側は急峻な崖が多く,道に迷ったら 命の危険もある。東側は,海岸の近くには平坦な地形になっているので,山の奥まで散策できる(大富林道と古見岳登山道)。この辺り(古見岳登山道)は 5 月から 7 月にかけて,毎日のようにベニボシカミキリの採集者が入っている。乾燥標本にして販売するのだろう。そろそろ天然記念物にして採集禁止にしてはどうか?

図 23.美田良(みたら)の田んぼ。この時は確か 8 月に行ったように記憶している。あぜ道を歩くと多くの生物を見ることができる。夜も危険はないので 長靴をはき,懐中電灯をもってあぜ道を歩くと,昼間見られない生物をたくさん観察することができる。田んぼの中にはオオヒライソガニがいる。

図 24.美田良の田んぼ道における生物観察。自然が好きな人には,西表島で自分の仕事とともに,生物の観察をお手伝いした。ただし,授業科目として実施すると大変なことになるので,本当に西表島の自然を見たい人たちに限定した観察である(単位なし)。宿泊は琉球大学熱帯圏研究センター・西表実験施設。教員は個室に入るので,実習中のストレスはあまりない。本当に自然を勉強したい人たちは,自己責任を自覚しており,実習もうまくこなす。

図 25.美田良の田んぼの畔を歩いて自然散策。畔には農薬が散布されているため,あぜ道には生物は少ない。・・・が,少し離れると多くの生物に出会える。サキシマハブは,雨のしとしと降る晩にはよく出てくるが,日中は見かけない。左の写真は西表島の典型的な自然林の景観。

図 26.自然観察。生物観察には,小型カメラ(Optio)を使った。Optio は何かの教育経費で 10 台ほど購入した。カメラは消耗品として購入した。海で使用したので壊れたり,紛失したりで購入後 5 年ですべて行方不明になった。 カメラは,備品としては管理が難しい。今は廃棄物の置き場で拾える。

図 27.美田良の田んぼにあった水草。四葉のクローバみたいなのが水面に浮いていた。こちらは被子植物だろう。水中にあるのは緑藻だが,種類は全然わからない。海には褐藻類や紅藻類が多いが,陸上は緑藻類が圧倒的に多い。何で 褐藻類や紅藻類は,陸上に進出して維管束植物になれなかったのか?

図 28.美田良の田んぼの脇にある林。イヌビワ,オオハマボウ,イワカワシジミ幼虫の食草である クチナシに混じってヒカゲヘゴが大きな葉を広げている。晴れた日には,道路や田んぼの脇の林縁部で多くの種類のチョウを見ることができる。このあたりだとナミエシロチョウ,イシガケチョウ,リュウキュウアサギマダラ,ツマムラサキマダラ,タイワンキチョウ,ヤエヤマウラナミジャノメ,セセリチョウ,コウトウシロシタセセリ,ルリウラナミシジミ,タイワンクロボシシジミが次から次に現れ,葉の上にとまる。ツマベニチョウもいる。西表島では海・陸問わず,豊かな自然環境を満喫できる。