
2024年1月29日(月)
1.Introduction
八重山諸島には,八重山層群(Yaéyama group)という中新統(Miocene Formation)が分布する。この中新統は,初めは八重山狭炭層(Yaéyama coal-bearing beds)と呼ばれたが,のちに八重山層群と呼ばれるようになった。八重山層群は,厚い砂岩と砂岩泥岩互層が主体となり,石炭層を挟む泥岩をともなう。
砂岩(sandstone)は,波によって海から運ばれた砂が堆積してできるのだろう。一方,泥岩(mudstone)は,陸上から運ばれてきた泥が堆積してできるのだろう。八重山層群のこのような特徴(厚い砂岩と砂岩泥岩互層)から見ると,この地層はユーラシア大陸東岸の沿岸・浅海域で堆積したと考えて間違いないように思われる。
ユーラシア大陸の起源は,ローラシア大陸(Laurasia)にある。古生代には,地球の北半球から南半球に広がる広大なパンゲア超大陸があった。パンゲアは,中生代の三畳紀(the Triassic)に入ると赤道を挟んで2つに分裂した。テチス海から北アメリカ大陸の南端に至る運河が形成され,北側はローラシア大陸,南側はゴンドワナ大陸となった。ジュラ紀(the Jurassic)に入ると,ローラシア大陸は,さらにユーラシア大陸と北アメリカ大陸に分裂し,両方の大陸の仕切りとなる運河は徐々に拡大し,両大陸は離れていった。
大陸の分裂と移動は,海産動物・陸上動物の双方に新しい生息環境を提供することになった。古生代の二畳紀(the Permian)と中生代の三畳紀(the Triassic)の境界に当たる時期には,大陸プレートの分裂と移動にともなって火山活動が活発になった。大規模な火山活動の影響は,地球上の大気や海洋環境を著しく悪化させ,結果として生物の大量絶滅(mass extinction)を引き起こした。火山活動の影響は特に海産動物が大きく,科(family)のレベルでは51%,属(genus)のレベルでは82%,種(species)のレベルでは95%が絶滅したという報告がある。特に沿岸域の海産動物は壊滅状態になっただろう。
しかしながら,二畳紀末の大量絶滅は,生物の進化にとってはむしろ好都合だったのかも知れない。大量絶滅に瀕したのは,古生代に生きた古いタイプの生物であった。海岸の浅瀬で,砂や石の上に体をさらして厚い外皮で捕食者から身を守っていただろう。動きは緩慢で,捕食者に対しても俊敏に逃げ回ることなどできなかったに違いない。
しかし古い生物でも,古生代という長い地質時代を生き抜いてきた訳だから,古生代の海洋環境によく適応していただろう。新しいタイプの生物が出現したとしても,それらに置き換わるまでには長い年月が必要だ。大量絶滅があって,古い生物が一気にいなくなってしまえば,置き換わりは迅速に進むはずである。・・・という訳で,三畳紀には新しいタイプの生物(十脚甲殻類で言えば,クルマエビ,ロブスター,イセエビ,ウチワエビ,アカザエビ,カニ類など)の進化が始まった。今(新生代第四紀)私たちの食卓に上がる十脚甲殻類の進化は,中生代に起きた大陸の分裂と移動と密接に関係している。
十脚甲殻類の進化は,より直接的には三畳紀以降に顕著になったサンゴ礁(coral reef)とも関係するだろう。古生代にもサンゴはあったが,礁(reef)は作らなかったのではなかろうか?大量絶滅の後に,生き残った刺胞動物(cnidarians)の中には,大陸や島の沿岸域の波の荒い磯に石灰質の硬い殻を作り,群体(colony)で生活する造礁サンゴ類が出現しただろう。大規模な造礁サンゴの群体は,個虫が生きているときだけではなく,死んだ後も石灰質の殻は残るので,サンゴ礁に住む多くの生物に隠れ場所を提供できる。私は,中生代における海産生物の大躍進(marine revolution)の基礎になったのは,造礁サンゴの出現だったのではないかと想像している。
十脚甲殻類で言えば,三畳紀の大陸沿岸の内湾(砂泥底)では,初めは卵をばらまくタイプのエビ(prawn)が進化した。パンゲア超大陸が分裂を始めると,海岸には陸上から大量の石や岩が転げ落ちてきただろう。それらは荒い波から身を守るための絶好の隠れ場所を提供したに違いない。イセエビやゾウリエビは,そんな環境で生命をつないできたのだろう。一方,三畳紀の内湾の砂泥底では,ロブスター(Homarus)の仲間が進化していた。ロブスターの進化と並行して,外洋に面する沿岸域には各所にサンゴ礁形成され,ロブスターの一部がサンゴ礁に移って生活を始めた。アナエビ類(Axiidea),ヤドカリやクダヒゲガニ等の異尾類(Amonura),カニ類の属する短尾類(Brachyura)は,ロブスターの祖先から進化したと思われる。
2.撮影と執筆の基本情報
<記事の執筆> 三枝誠行(生物多様性研究・教育プロジェクト常任理事)。
3.参考文献
・Brand, U., R. Posenato, R. Came, H. Affek, L. Angiolini, K. Azmy, and E. Farabegoli (2012) The end‒Permian mass extinction: a rapid volcanic CO2 and CH4‐climatic catastrophe. Chem. Geol. 322–323: 121–144.
・Erwin, D.H. (1990) The end-Permian mass extinction. Annu. Rev. Ecol. Syst. 21: 69‒91.
・Erwin, D.H., S.A. Bowring, and J. Yugan (2002) End-Permian mass extinctions: A review. Geological Society of America Special Paper 356: 363–383.
・Jablonski, D. (1991) Extinctions: a paleontological perspective. Science 253: 754‒757.
・神谷厚昭(2001)西表島の地形と地質‒露頭の紹介を中心として。西表島総合調査報告書‒自然・考古・歴史・民俗・美術工芸。沖縄県立博物館。
・木村政昭(1996)琉球弧の第四紀古地理。地学雑誌 105: 259‒285。
・小竹信宏・亀尾浩司・奈良正和(2013)沖縄県西表島の中部中新統西表層最上部の地質年代と堆積環境。地質学雑誌 119: 701‒713.
・Saitoh, Y., and F. Masuda (2004) Miocene sandstone of ‘continental’ origin on Iriomote Island, southwest Ryukyu Arc, Eastern Asia. J. Asian Earth Sci. 24: 137–144.
・平朝彦(1990)日本列島の誕生。岩波新書。
・Vermeij, G.J. (1977) The Mesozoic marine revolution: evidence from snails, predators and grazers. Paleobiol. 3: 245-258.
・Vermeij, G.J. 1987. Evolution and Escalation. Princeton University Press.
・Futuyma, D.J. 1998. Evolutionary Biology. Third edition. Sinauer Associates Publishers.
・William, B.B., and H.W. Warren (1960) A survey of the resources and economic potential of Iriomote Island. Rept. Geol. Min. Res. Stanford Res. Inst. VI, 1–10.
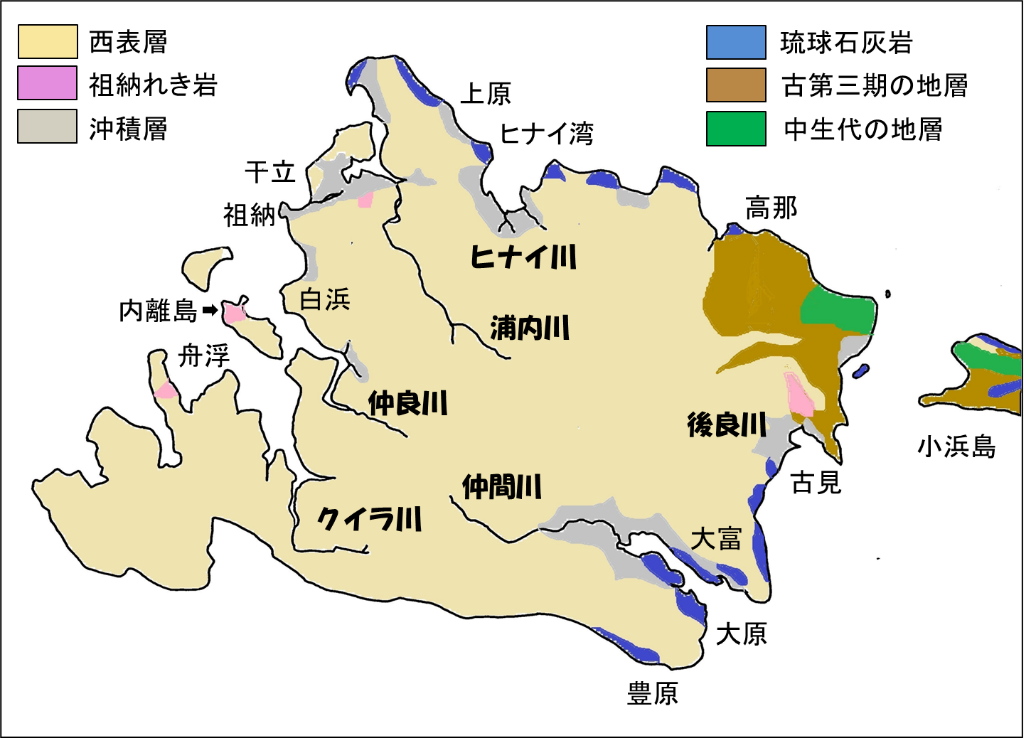
図 1.西表島の地層。最上層は八重山層で覆われている。八重山層の厚さは不明だが,最大 700m の厚さがあるとどこかに書かれていた。
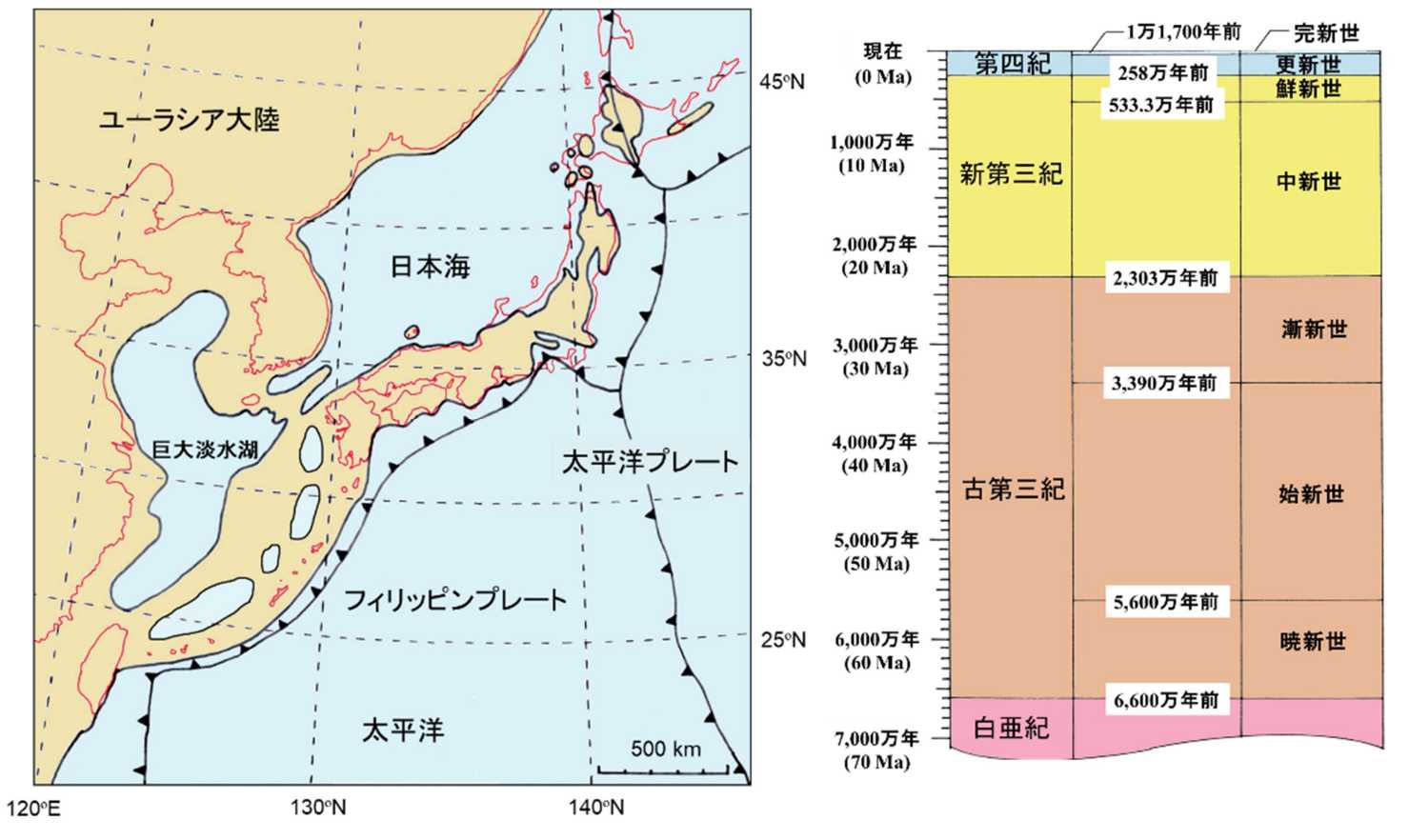
図 2.(左)500 万年前の日本列島周辺の地形。(右)新生代の地質年代区分。500 万年前というと,新生代第三紀の中新世と鮮新世の境界に当たる。日本列島は現在の地形に近づきつつある。ユーラシア大陸と琉球弧の間には広大な淡水湖が広がっていて,中国大陸ではコイ科を中心として種の多様化が進んでいたかと推察される。琉球弧の周辺にも淡水湖がいくつかあったが,どんな生態系があったのか,想像しにくい。日本海の方は,北海道と沿海州(ロシア)の間には広い海峡があり,海水の塩分濃度は割と現在に近かったかもしれない。日本海に生息するベニズワイは,500 万年ほど前から日本海に入り込んでいたかもしれない。琉球弧はユーラシア大陸と 2 か所で陸続きになっていて,淡水湖の北と南から生物が分布を広げただろう。
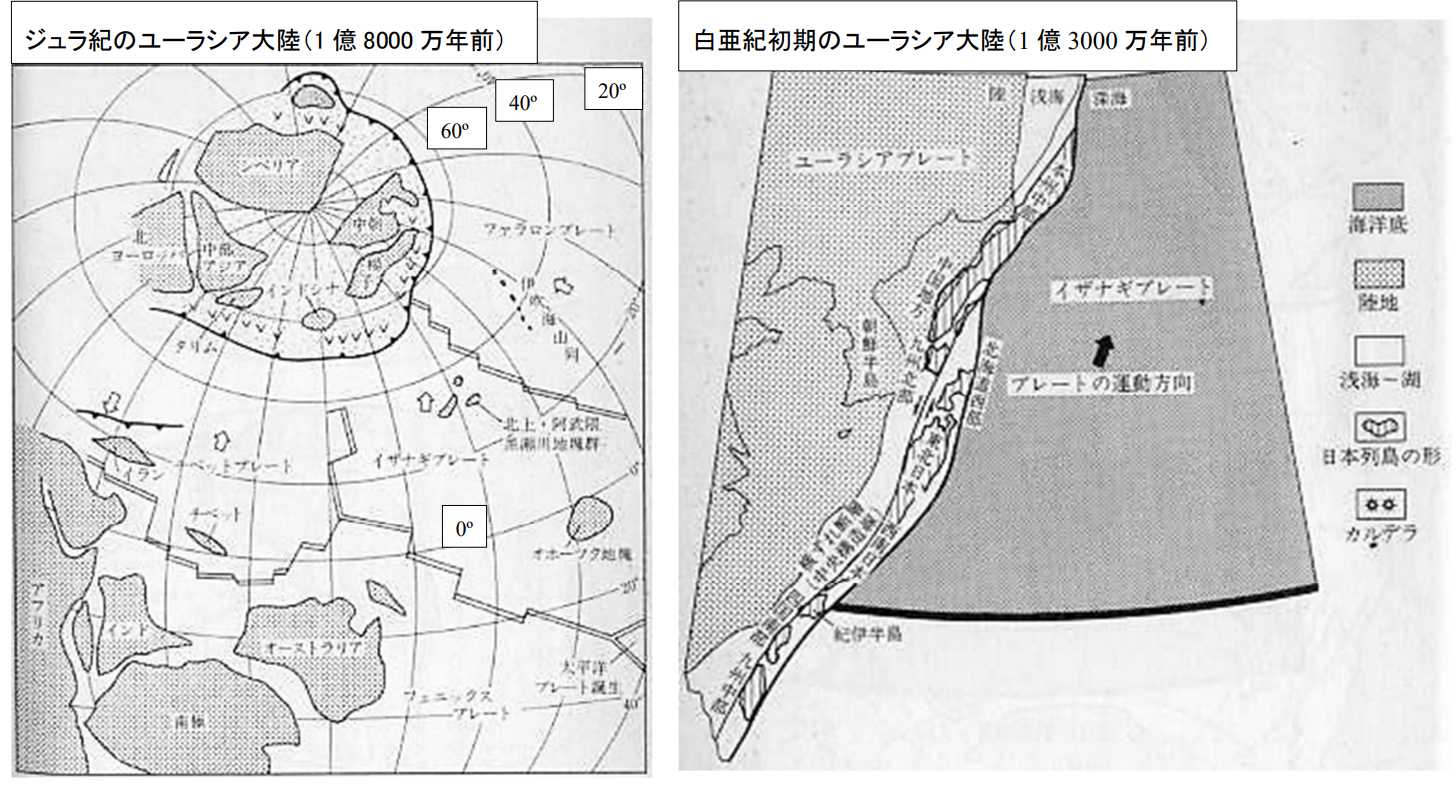
図 3(左)。ジュラ紀初期のユーラシア大陸。平朝彦「日本列島の誕生」③から転写。ローラシア大陸はジュラ紀になってから,ユーラシア大陸と北アメリカ大陸に分裂した。問題は,三畳紀からジュラ紀にかけてのローラシア大陸の位置である。平朝彦「日本列島の誕生」では,ローラシア大陸(ユーラシア大陸+北アメリカ)は北極圏にまとまっている。ジュラ紀に地球上に出現した始祖鳥(Archaeopteryx)は羽毛をまとっていたという証拠がある。羽毛があるということは,寒さに対する適応なので,ジュラ紀のユーラシア大陸が高緯度にあったと考えることは可能である。しかし,いくら何でも,ローラシア大陸全体がほとんど北極圏に入っているというのはちょっと考えにくい。
図 3(右)。白亜紀の横ずれ運動。平朝彦「日本列島の誕生」④から転写。白亜紀のユーラシア大陸が地球上のどの位置にあるかは不明であるが,大陸の縁を北海道南部,東北日本,西南日本と紀伊半島,四国南部,九州中部がそれぞれ島となって,大陸の東端を次々と高緯度に移動している。白亜紀にはまだ琉球弧と言える地形(大陸棚の浅瀬)が存在していない。ただ,無理に理屈をつければ,「存在する」という結論にはできる。
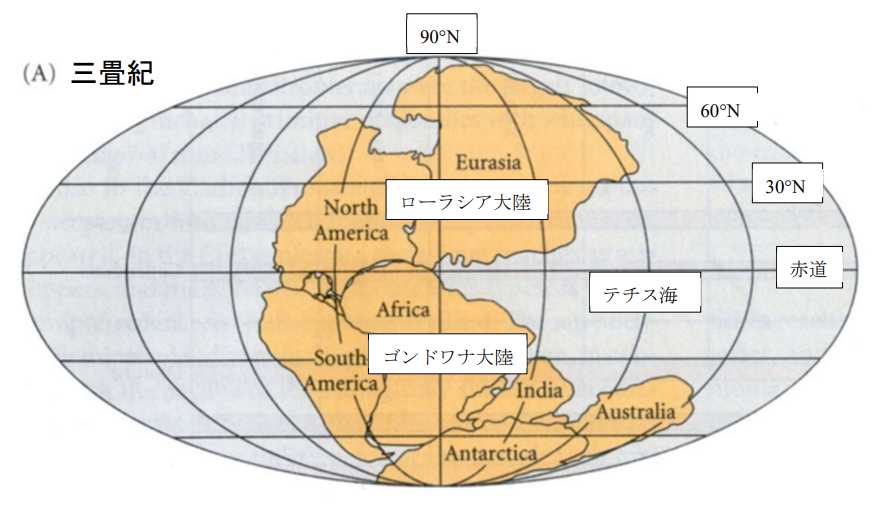
図4.三畳紀の大陸。Futuyma(1998)から転写。
古生代にはパンゲアという超大陸が存在していたが,中生代に入り,パンゲアのあちこちに裂け目が入り大陸の分裂が始まった。北半球では,まず分裂したのが,ローラシア大陸とゴンドワナ大陸である。
三畳紀には,ユーラシア大陸は赤道から北極圏におよび,面積は現在とほぼ同じだったと思われる。北アメリカは「大陸」と呼んでよいのか微妙なところだと思うが,赤道以南(10ºS ぐらい)から北緯 60º 未満に位置していた。
ゴンドワナ大陸の方は,アフリカ大陸,南アメリカ,インド,オーストラリア,それに南極大陸への分裂が始まった。
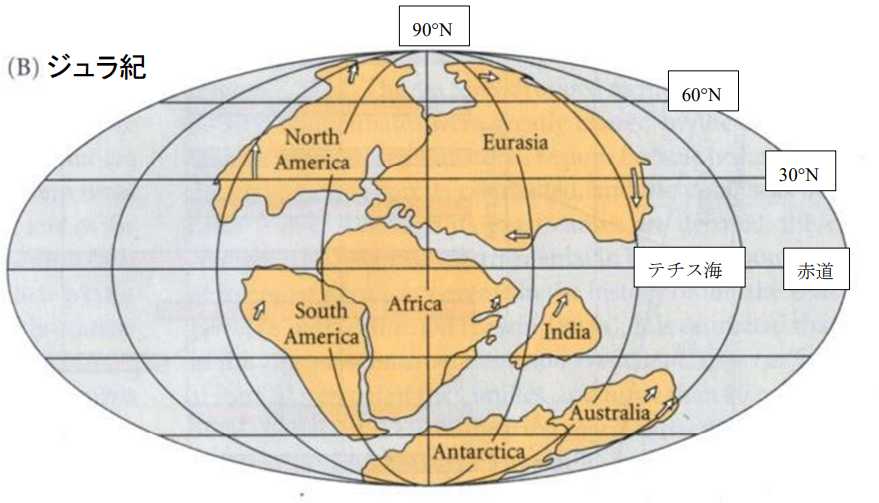
図 5.ジュラ紀における分裂後の大陸移動。Futuyma(1998)から転写。
ユーラシア大陸は,現在の中央アジアを中心として時計回り(右回り)に回転している。現在の日本列島や琉球弧に対応するユーラシア大陸の沿岸域は,現在よりもずっと高緯度にあったことが推察される。一方,北アメリカの方は,どんどん北上を続けたのかもしれない。ジュラ紀には,ユーラシア大陸と北アメリカはまだ近接した位置関係なったのかもしれない。
アフリカ大陸と南アメリカは,現在よりも少し北に位置していた可能性がある。オーストラリアはまだ南極大陸とくっついていたが,インド亜大陸は,アフリカ大陸から大きく離れてユーラシア大陸に向けて移動している。
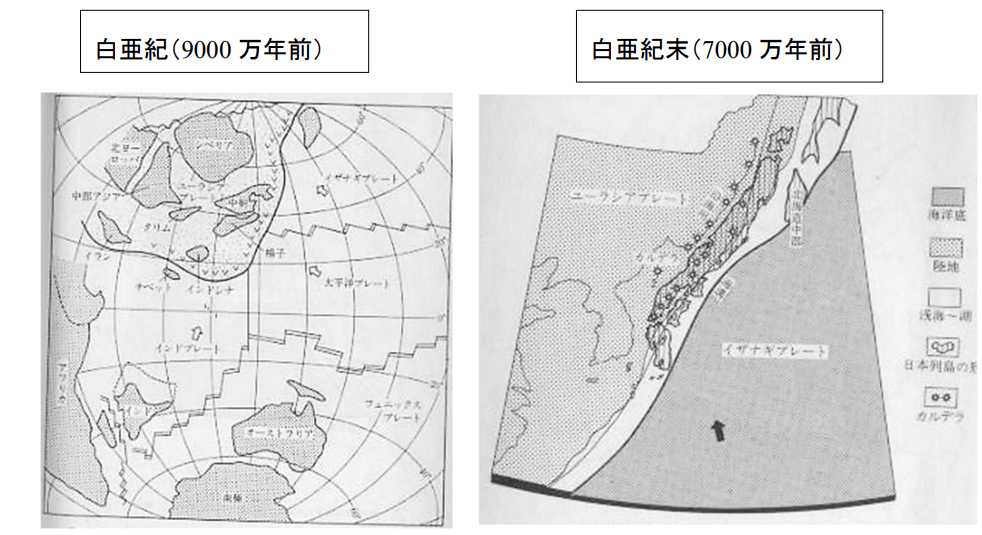
図 6(左)。白亜紀のユーラシア大陸の位置(9000 万年前)。平朝彦「日本列島の誕生」⑤から転写。白亜紀になるとユーラシア大陸は低緯度に移動しているが,北アメリカの方は不明。ユーラシア大陸の東部には,イザナギプレートと太平洋プレートがともに北西に向かって移動している。ユーラシア大陸は,白亜紀には時計回りに回転しているか,低緯度に向かって南進している。どちらにして
も,日本列島がどの場所にあるのか特定するのは難しい。
図 6(右)。白亜紀のユーラシア大陸東端の地形(7000 万年前)。平朝彦「日本列島の誕生」⑥から転写。日本列島が,ユーラシア大陸のどの位置にあったかは不明。
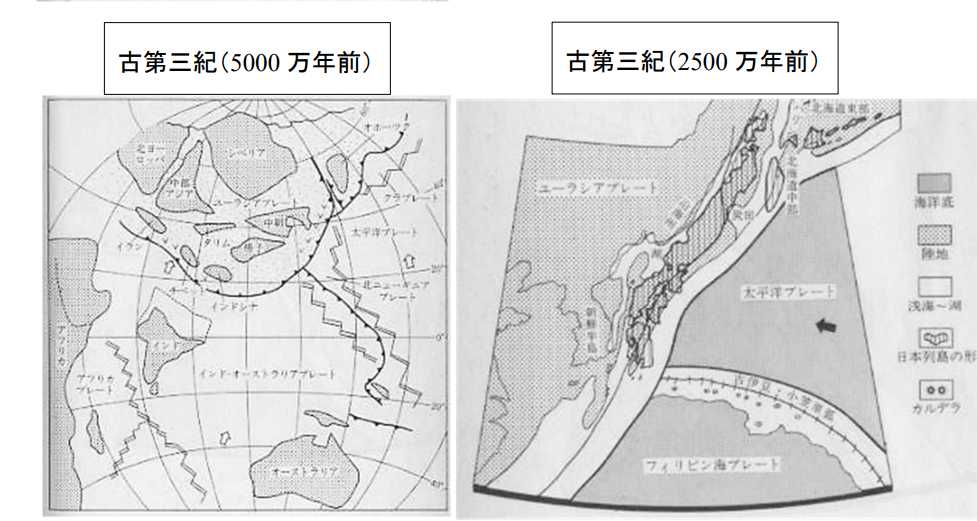
図 7(左)。新生代古第三紀のユーラシア大陸(5000 万年前)。平朝彦「日本列島の誕生」⑦から転写。新生代に入ると,ユーラシア大陸(点刻した区域)は現在とほぼ同じ緯度にある。ユーラシア大陸の東側には太平洋プレートと北ニュ
ーギニアプレートがせめぎあっているが,フィリッピン海プレートは出現していない。
図 7(右)。新生代古第三紀のユーラシア大陸(2500 万年前)。平朝彦「日本列島の誕生」⑧から転写。2500 万年前(25 Mya)になると,フィリッピンプレートが出現し,古伊豆・小笠原弧が出現している。琉球弧の位置は確定していたと思うが,島はまだ現れていない。
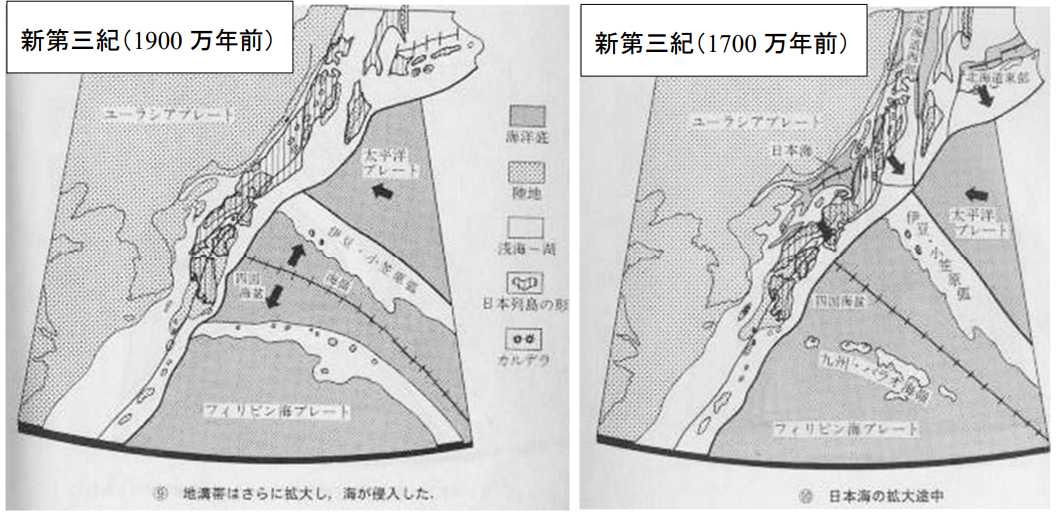
図8(左)。新生代新第三紀の日本列島(1900 万年前)。平朝彦「日本列島の誕生」⑨から転写。1900 万年前だと,日本海はでき始めたころでまだ浅い。琉球弧では島が出現しているが,奄美大島や沖縄本島,周辺の島々かは不明。伊豆島(現在の伊豆半島)は,まだ硫黄島から小笠原方面にあったと思われる。
図 8(右)。新第三期の日本列島(1700万年前)。平朝彦「日本列島の誕生」⑩から転写。日本海が拡大している。「日本海」の矢印は深いところ(海洋底)を示す。大陸を示す点刻と紛らわしいので注意。
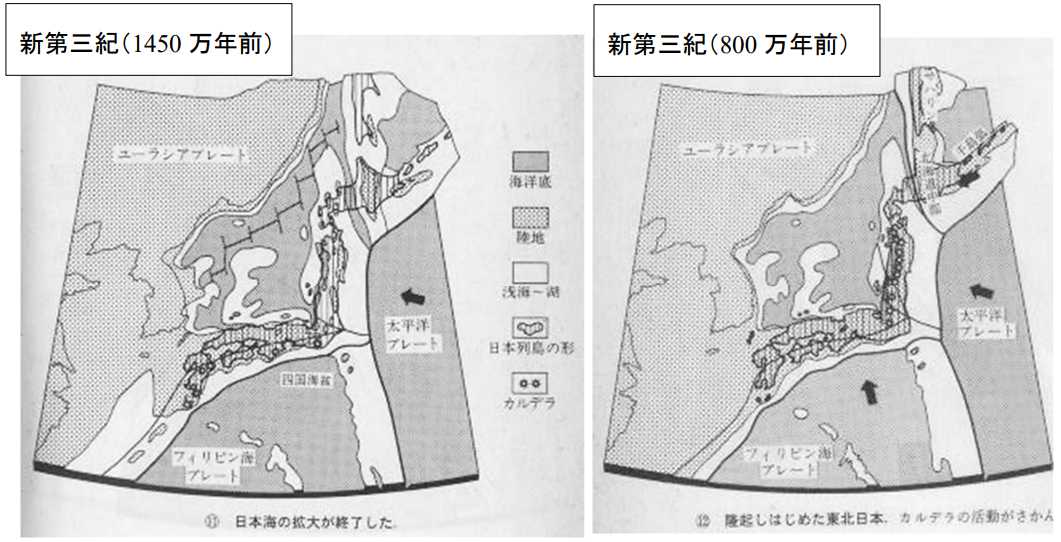
図 9。新生代新第三紀の日本列島。左が1450 万年前,右が 800 万年前。平朝彦
「日本列島の誕生」⑪と⑫から転写。
日本海が現在とほぼ同じ形になったころの日本列島。九州沿岸とユーラシア大陸は地続き。日本海は,大和堆は比較的浅いが,周辺は深くなっている。本州は,能登半島のところで北と西に直角に折れ曲がっている。
琉球弧はユーラシア大陸の縁に出現しており,大小さまざまな大きさの島が点在していただろう。八重山諸島の地形は,このころから隆起をはじめ,西表島は海面から顔を出したのではないか。
800 万年前には,琉球弧はユーラシア大陸の縁にあって陸続きになっている。
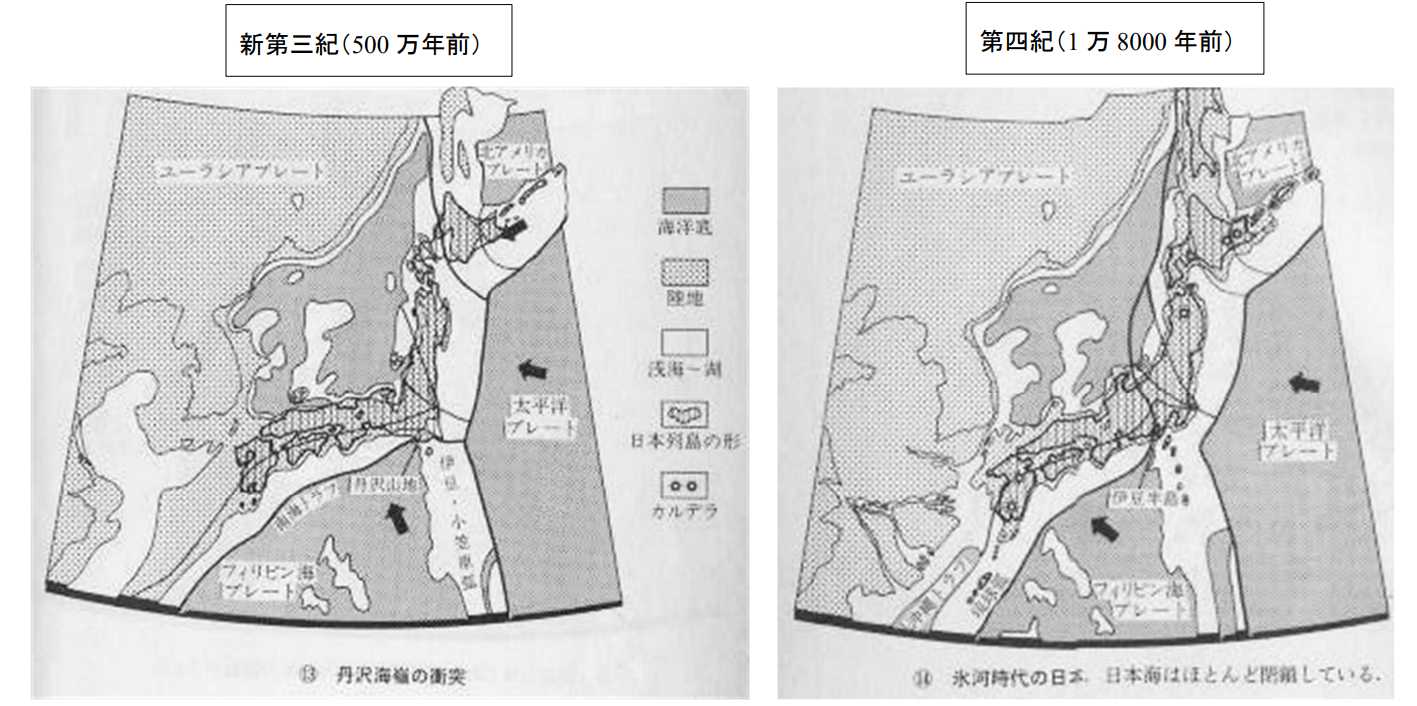
図 10(左)。新生代新第三紀(500 万年前)の日本列島。平朝彦「日本列島の誕生」⑬から転写。日本海は宗谷海峡でオホーツク海とつながっているが,九州側では陸封されているため,日本海の塩分濃度は極めて低かっただろう。図 2 を見て,ベニズワイは 500 万年ほど前に日本海に侵入したと書いたが,日本海が 10‰(パーミル)程度の塩分濃度だったとしたら,とても生きて行けなかっただろう。一方,九州の西方にあった淡水湖ではコイ科(淡水魚)の種分化が進んだかもしれない。
図 10(右)。新生代第四紀(1 万 8000 年前)の日本列島。平朝彦「日本列島の誕生」⑭から転写。図 10 では,伊豆半島は硫黄島の付近から移動したのではなく現在の位置で隆起した可能性がある。どっちの仮説が正しいのだろうか?九州と大陸の間が海で仕切られたのが氷河期以降になるので,ベニズワイは氷河期が終わったころ(1 万年前ぐらい)に日本海に入ってきたのかもしれない。1 万年前というと,日本列島は旧石器時代が終わり,縄文時代に入っている。ベニズワイは日本人の歴史で言えば,縄文時代になってから日本海に定着した可能性が高い。
1 万 8000 年前には,琉球弧の西側に沖縄トラフが形成されている。次のページの図 11(左上)には,琉球弧に沿って広大な淡水湖が見られる。淡水湖は沖縄トラフ形成の一環として生じたものだろう。
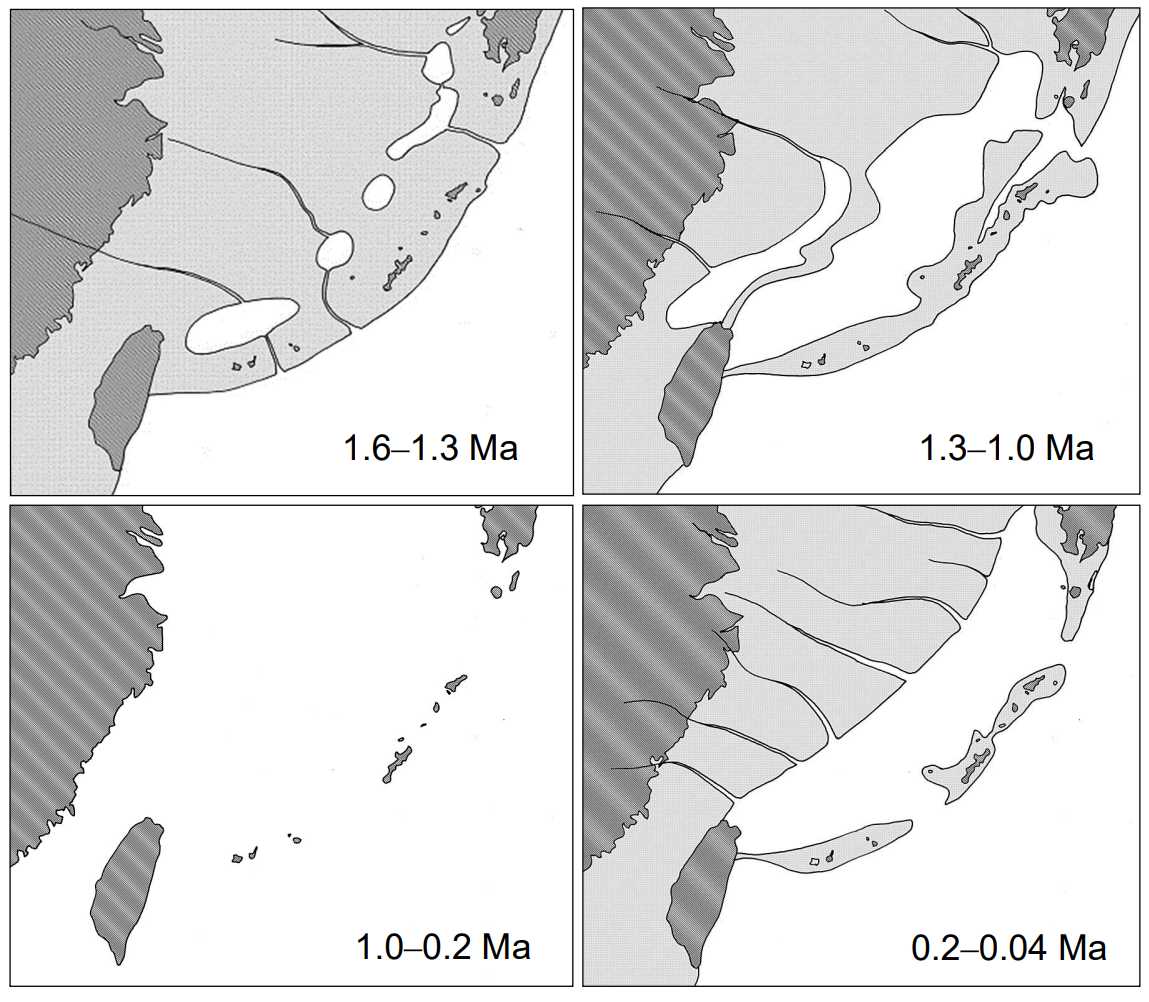
図 11.第三紀末期から第四紀にかけての琉球弧の古地理。木村政昭(1996)から図を描き直した。
(左上)160 万年前から 130 万年前のユーラシア大陸東岸。琉球弧はユーラシア大陸と地続きになった。台湾から九州にかけては広大な陸地が広がっている。淡水湖が沖縄トラフになってゆくと思われる。
(右上)130 万年前から 100 万年前。ユーラシア大陸沿いの広大な淡水湖(汽水湖)は奄美大島と屋久島の間で太平洋とつながっている。
(左下)100 万年前から 20 万年前の琉球弧。島々の地形は現在とよく似ている。この時期は琉球弧で絶滅した動物や植物が多いのではないか?
(右下)20 万年前から 4 万年前の琉球弧。台湾とユーラシア大陸は地続き。奄美諸島と沖縄本島は地続き。西表島から宮古島に至る広大な島は,台湾と地続きになっている。
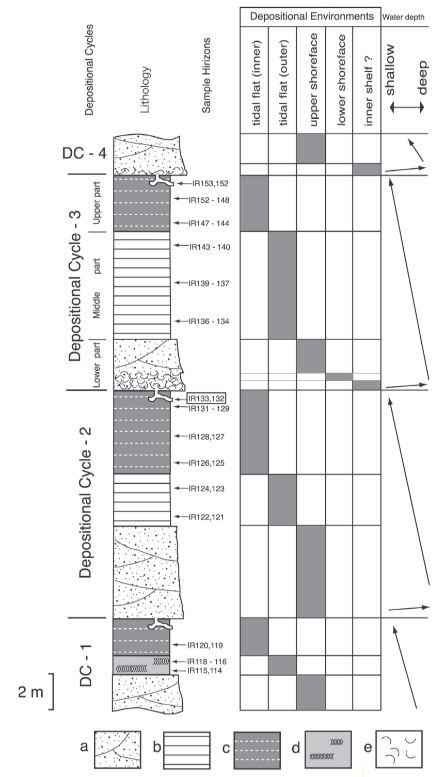

図 12.西表島の海岸に露出している地層の区分。小竹ら(2013)の論文から転載。私が日本語で解説するよりも,英語の説明を読んだ方が理解しやすいかもしれないので,図の説明の原文も掲載した。 西表島の隆起や沈降にともなって,島の周囲にある海岸や河口の地形と標高は異なる。地形や海抜が変化すれば,それらに応じて堆積する土壌も異なる。
堆積環境の inner tidal flat は,河口域とマングローブに堆積する地層を示す。河口やマングローブには,陸上から運ばれてきた泥(mud)が堆積する。図 12(Fig. 3)で言えば,各堆積サイクルの中の地層 c や地層 d が該当する。
Outer tidal-flat は,外洋に面する内湾であろう。湾になると,海から波に運ばれて砂(sand)が堆積する。外洋に面する内湾だと,泥と砂の入り混じった土壌(地層 b)が堆積する。
Upper shoreface は,外洋に面する湾の砂浜海岸だろう。砂岩の層(地層 a)ができる。Lower shoreface は,外洋に面する内湾のやや深いところで,堆積する砂の粒度も大きくなり,貝殻や造礁サンゴのかけらも含まれるだろう(地層 a)。
Inner shelf というと,大陸棚の内側ということになるが,深さが100m に達するぐらいの海底だろうか?貝殻を多く含む。

図 13.西表島西海岸の露出した地層(西表層)。砂岩の層(図 12 の a)が中心で,その中に泥岩の薄い層(図 12 の c)が挿入されている感じである。
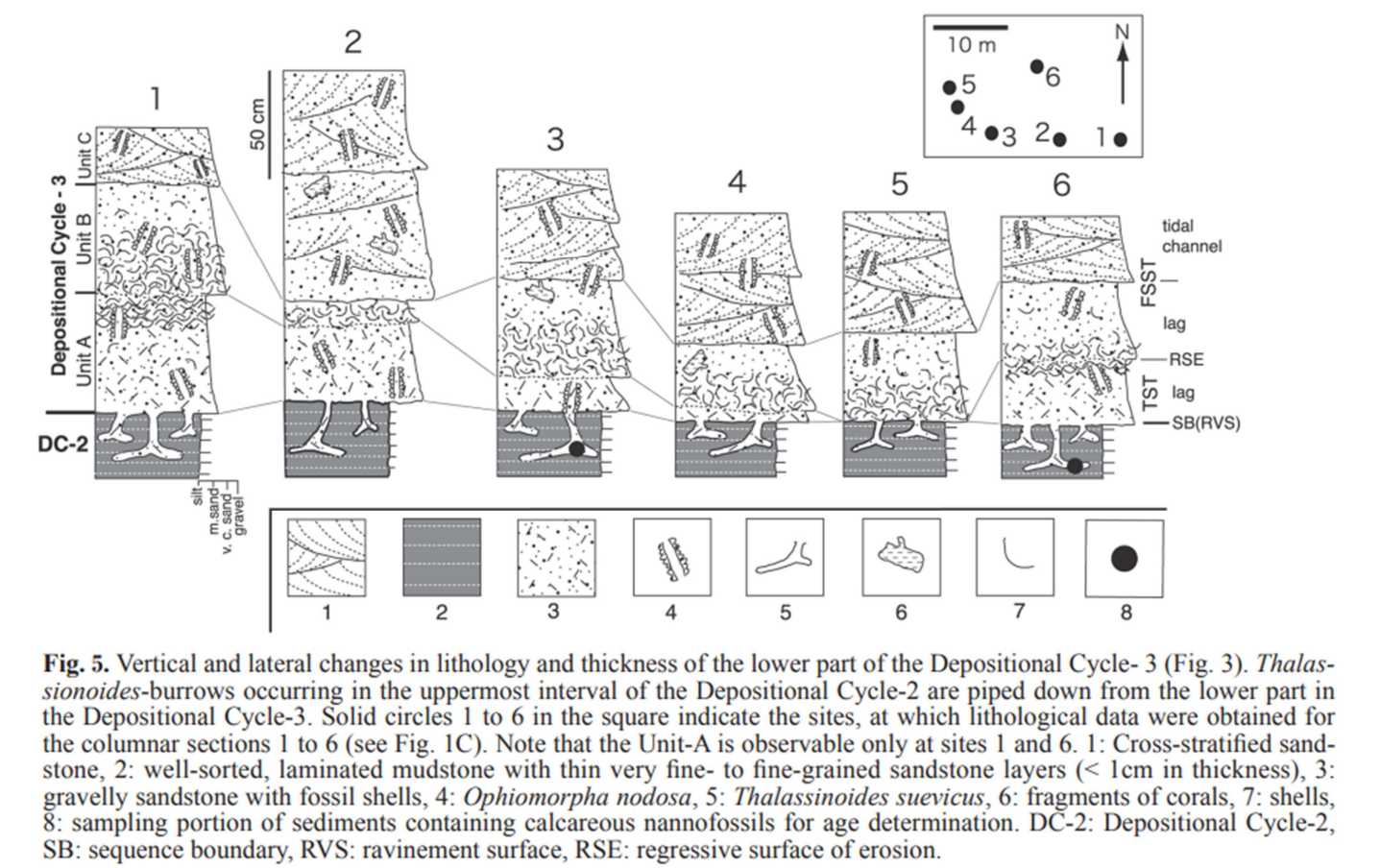
図 14.西表島の堆積層の中に含まれる 2 種類の生痕化石。図(Fig. 5)は小竹ら(2013)の論文から転載。
堆積サイクル 3 で言えば,地層 a (図 13)に対応するのは,図 14 では地層 1 と地層 3 の混合層になるのだろう。どちらも砂を主体とし,貝殻の化石を含む。この層(地層 1 と地層 3 の混合)の中に,生痕化石 Ophiomorpha nodosa が発見される。一方,堆積サイクル 2(図 13)の地層 2(図 14)は,薄い砂岩の層に含まれた泥岩層であり,生痕化石 Thalassinoides suevicus が発見される。どちらの生痕化石もどんな種類か不明。

図 15.浦内川河口の干潟(tidal flat)。正面に見える背の低いヒルギは,和名がヒルギダマシ,学名は Avicennia marina。その右にある 3 本のヒルギは,和名がメヒルギ,学名は Kandelia obovata。その向こうに一列に並んでいるのはヤエヤマヒルギ(Rhizophora stylosa)。誰かが植林をしたか?

図 16.浦内川河口の干潟を歩く調査隊。このあたりに生えているのはすべてヤエヤマヒルギ(Rhizophora stylosa)。底質は硬く,ぬかるんでいるところが少ない。干潟の上を歩いても足を取られない。水たまりには猛毒のツムギハゼが普通に泳いでいる。個体数も多い。

図 17.浦内川河口の干潟の底質。海から運ばれた砂と、陸上から運ばれた泥が入り混じっている。このあたりの汽水は富栄養化しているだろうが,泥の色や海水の色を見る限り,酸素は泥砂の底質に行き渡っているように見える。干潟生態で注目すべき 3 つの重要な課題は,堆積した泥砂の粒度(grain size),海水(汽水)の富栄養化(eutrophication),重金属における海洋汚染(heavy-metal contamination)が干潟の生物の生態・行動・生殖にどんな影響を及ぼすのかを明らかにすることである。しかし,全国の理学部にある研究室の中で,こんなことをやっているのは私たちだけだっただろう。
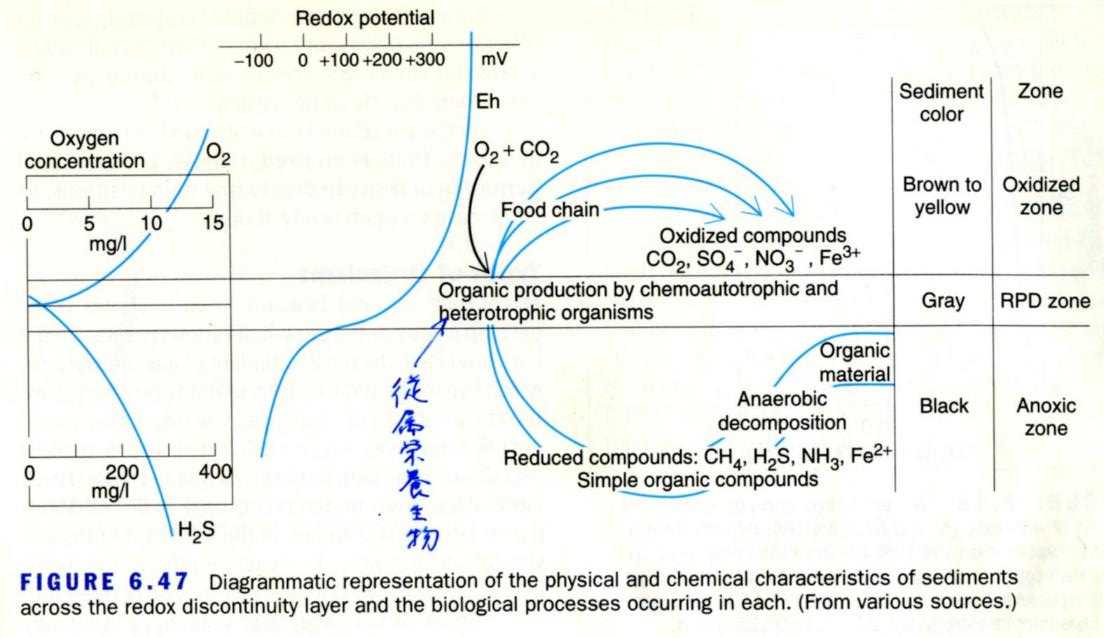
図 18.干潟の堆積物(sediments)の物理・化学的性質。Nybakken, J.W. (2001) Fig. 6. 47 を転写。砂泥の色彩に注目していただきたい。茶色から黄色の層には十分な酸素が含まれている。灰色は酸素濃度が低下している(RPD ゾーン)。黒くなると,還元化された硫化物のために,浜は硫黄のにおいがする。好気性の生物は住めなくなり,嫌気性の細菌が繁殖する。
<参考文献>
Nybakken, J.W. 2001. Marine Biology, An Ecological Approach. Fifth edition. Benjamin Cummings, San Francisco, CA.
Yamasaki, M., T. Nanri, S. Taguchi, Y. Takada, and M. Saigusa (2010) Latitudinal and local variations of the life history characteristics of the thalassinidean
decapod, Upogebia yokoyai: A hypothesis based on trophic conditions. Est. Coast. Shelf. Sci. 87: 346‒356.
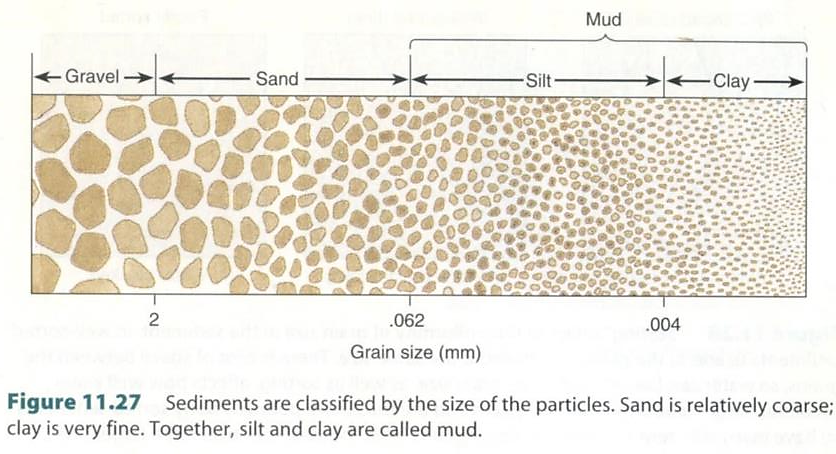
図 19.干潟に堆積する砂や泥の粒径分布。Castro, P., and M.E. Huber(2005)Figure 11.27 を転写。
干潟の堆積物は,主には砂と泥である。砂は大体径が 620 µm(0.62 mm)以上の荒い粒子からできており,泥の方は,620 µm 以下の細かい粒子からできている。粒径が 2 mm 以上になると gravel (礫岩)になる。一方,泥の方は粒度の径が 40 µm(0.004 mm)以下を粘土(clay)と呼び,0.04 mm と 0.62 mm の間にある粒子をシルト(silt)と呼んでいる。このように,地質学の分野では砂と泥の間には厳密な区別があるが,生態学的な研究をやるならば,「砂」と「泥」が区別できれば問題ないだろう。
生態学研究では,むしろ gravel の粒径以上の大きさのサンゴ塊とか砂泥岩のブロックの方が重要である。例えば熱帯域・亜熱帯域には,サンゴ塊が強い波に砕かれて堆積している海岸が多い。海岸の底質が砂地だと,穴を掘って棲む生物は少ないが,底質に泥が多く混じると,多くの生物が穴を掘っている。死んだサンゴ塊も,でこぼこした硬い石灰質の隙間に多様な生物が入り込んでいる。
死んだサンゴ塊の上に形成された干潟の場合には,砕かれたサンゴ塊を何と呼ぶのか?coral rubble? coral gravel? coral pebble?どれもこれが正しいという表現はない。私は coral rubbles をよく使うが,一番良い表現かはわからない。
干潟においては,生物がどんな生息環境に置かれているかが重要である。日本の場合には,どの研究者も判で押したように,種内関係や種間関係とか,あるいはどんな種かを議論しているが,その前に生息環境の物理・化学的要因を正確に把握しておくことが重要ではないかと思う。日本の科学者は視野が狭すぎ,全体像を把握することが苦手な人が多く,権威主義的な学問が横行する原因になっている。

図 20.干潟に堆積する泥や砂の粒径と水の通過性との関係。Castro, P., and M.E. Huber(2005)Marine Biology. McGraw Hill Higher Education のFigure 11.27(図 19)と Figure 11.28 を転写。
(a) 堆積した土壌の粒径が大きく,また粒径もそろっている(well-sorted)場合には,水の通過が早い。
(b) 堆積した土壌の粒径が小さく,粒径がそろっている場合には,(a)と比べて,侵入した水が土壌の層を通過する速度が遅くなる。
(c) 堆積した土壌の粒径がまちまちで,粒度の大きな砂から非常に粒度の小さい泥が混じっている場合には,水の浸透速度が著しく低下する。瀬戸内海の湾に堆積したヘドロは,このケースで,土壌を通過するのに時間がかかるために,水に溶けた酸素が酸化によって奪われてしまうことが大きな環境問題につながっている。
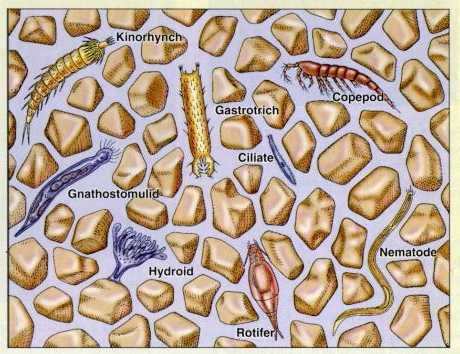
図 21.堆積した土壌の中に棲む生物群集(meiofauna)。Castro, P., and M.E. Huber(2005)Marine Biology の 273 ページの Figure(番号はない)を転写。堆積した土壌の粒径が大きい場合には(例えば,粒径が 5mm 前後)小石と他の小石の間に空間ができる。粒度の細かい泥が水に混じって入ってくるような土壌であれば,小石と小石の隙間には多くの微小生物(間隙性動物)が見られる。
copepod は甲殻綱の橈脚目。kinorhynch は袋形動物門の動吻綱,gastotrich は胃繊毛門,cilliate は原生生物門の繊毛虫綱,hydroid は刺胞動物門のヒドロ虫綱,rotifer は輪形動物門,nematode は袋形動物門の線虫綱に属する。体長は 0.5mm から 2~3mm の動物である。先カンブリア紀から古生代にかけて繁栄したが,すでに大半は絶滅しただろう。
間隙性動物を調べるには,フィールド図鑑は役立たない。下記の文献を読んで,原生生物門や袋形動物門のところを見たらよい。
<参考文献>
Ferl R.J., and R.A. Wallace. 1996. Biology. 英語で読んだら勉強になる良い本である。
山田・西田・丸山 1981. 進化系統学。日本に新しい系統分類学の風を吹き込んだ名著。
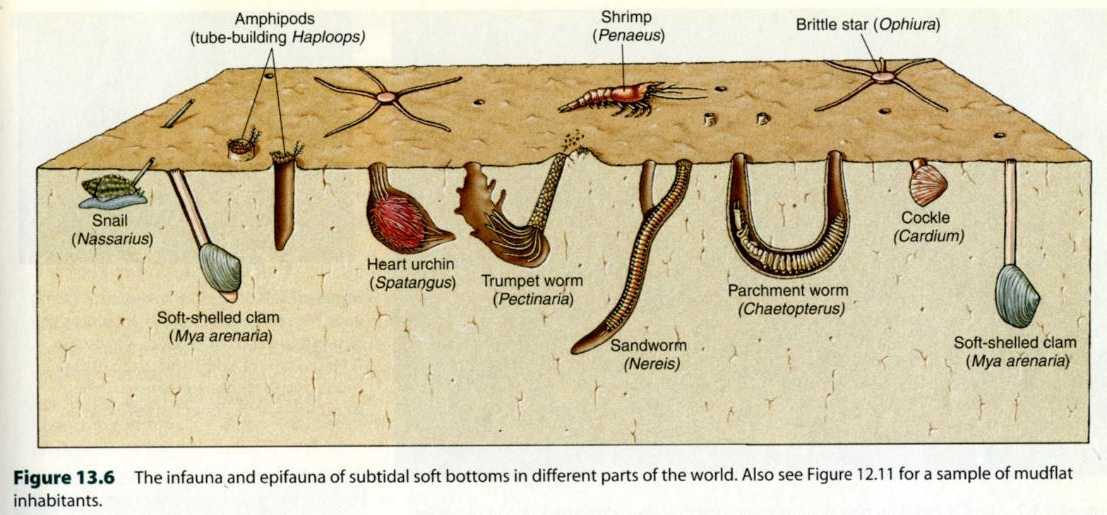
図 22.干潟の砂泥底で生活する大型の無脊椎動物(invertebrates)は,穴を掘って生活する。土中に酸素がなくても,鰓を使って海水を循環させれば,必要な酸素は確保できる。穴を掘るのは大型の動物で,環形動物・軟体動物・節足動物が多いだろう。Amphipods(左上)は Annelids の間違い。
干潟の生物は泥を食べて生きているのか?
中学校や高等学校の生物の教科書には,干潟の砂泥底で生活する動物は,干潟の泥をすくって口に入れ,砂の中から有機物をこしとって食べる「ろ過摂食」(filter feeding)と書かれているかもしれない。環境教育学や海洋生物学の教科書でもそんな表現が一般的と思われる。
もし試験にシオマネキ等の干潟の動物は,泥を食べているかという問題が出たら,正しい答えとしてあなたはどの解答を選択するだろうか?「食べていない。なぜならば,シオマネキはろ過摂食だから。」という解答を選んだら,多分正解になると思う。
しかし,上記の解答は実は不正解である。確かにシオマネキはすべてろ過摂食だから良いのであるが,干潟には効率は著しく悪いが,泥を食べてしまう動物もたくさんいる。考える機会を与えず,知識だけで正解を決めるシステムは悪い側面も持っている。寄生と共生の問題も似たところがある。

図 23.マングローブの中の砂泥底に棲む動物の調査。
泥の中に穴を掘る動物を採集するにはベイト・ポンプが役に立つ。
海洋に流出した汚染物質が,干潟に棲む生物の生態・行動・生理や内分泌に及ぼす影響については,多方面からの研究が進められている。悪影響や異常を記述した原著論文や総説は数多く出版されている。しかし,異常とみなすにはコントロール,つまり正常な生物の生活については全く調べようとしない。だから,何が本当に異常なのかは,正常な生活をしている生物を見て判断する必要がある。
そういうことで,日本列島の多くの干潟で,どんな生物がどんな生活をしているのかを知るために「干潟プロジェクト」が始まった。主な調査場所は瀬戸内海の干潟で,西表島で行う必要性はなかったが, 西表島の干潟について詳しく知りたいので研究に加えた。
干潟プロジェクトは,ある学生が担当した。この学生は,東京都立工専から 3 年次編入で研究室に配属された。数学と理科(物理と化学)は素晴しい成績を収めていたが,国語に関しては,からきしダメだった。よく仕事はしたが,英語でも日本語でも文章が書けず,博士号を取得するまでに,私の方は大変な苦労をすることになった。
<参考文献>
・B.J. Kang, B.J., T. Nanri, J.M. Lee, H. Saito, C.-H. Han, M. Hatakeyama, and M. Saigusa (2008) Vitellogenesis in both sexes of gonochoristic mud shrimp, Upogebia major (Crustacea): analyses of vitellogenin gene expression and vitellogenin processing. Comp. Biochem. Physiol. Part B:
Biochem. Mol. Biol. 149: 589-59.
・Nanri, T., M. Fukushige, J.P. Ubaldo, B-J. Kang, N. Masunari, Y. Takada, M. Hatakeyama, and M. Saigusa (2011) currence of abnormal sexual dimorphic structures in the gonochoristic crustacean, Upogebia major (Thalassinidea: Decapoda), inhabiting mud tidal flats in Japan. J. Mar. Biol. Ass. U.K. 91: 1049 – 1057.
・Ubaldo, J.P., T. Nanri, Y. Takada, and M. Saigusa (2014) Prevalence and patterns of infection by the epicaridean parasite, Gyge ovalis and the emergence of intersex in the estuarine mud shrimp, Upogebia major. J. Mar. Biol. Ass. U.K. 94: 557‒566.

図 24.浦内川の干潟。干潟には小さなマウンド(泥の塚)が無数にある。棲んでいるのは一種類のスナモグリ(Callianassa bouverii)だろう。

図 25.浦内川の干潟に生息する穴掘り生物。干潟にはカニやエビは多いが,穴を掘って棲む十脚甲殻類は少ない。上が Calianassa bouverii(スナモグリ)。下が Upogebia iriomotensis か U. neogenii(アナジャコ)。U. iriomotensis と U. neogenii は形態的には区別できない。遺伝子解析では容易に区別できる。

図 26。泥を食べる干潟生物。うんちの太さからして相当大きな環形動物だろう。ミミズもそうだが,泥や砂を食べると栄養的にはものすごく効率が悪いだろう。節足動物になると,泥や砂をすくって食べるが,口器(顎脚)で有機物と砂を営利分けて食べるようになった。摂食方法も進化した。

図 27.浦内川の干潟。干潟ができるのは干潮時に限られる。水たまりには小さなハゼが多い。正面にあるのはヤエヤマヒルギ。右側の 2 本はメヒルギ。ずっと奥には祖納岳の鉄塔が見える。ときどき干潟の上をナミエシロチョウやアオスジアゲハが横切る。何百年も変わっていない風景だろう。

図 28.ミナミコメツキガニの集団。ミナミコメツキガニは干潮時に干潟ができると集団で歩き,盛んに鋏脚で泥をすくって食べている。口の中に入れるのは有機物で混じった砂は顎脚を使って外に捨てるのだと思う。歩いた周辺にはカニが作った小さな砂団子がいっぱい散らばっている。

図 29.ミナミコメツキガニの大集団。西表島ではどこの河口でも干潮時になると,こんな集団が普通に見られる。圧巻の風景である。

図 30.タテジマユムシ(環形動物)。干潟の泥に潜り,白い触手(と言ってよいか?)を,本体(赤い袋)が入っている穴を中心に 50 ㎝四方に伸ばし,触手の先端にある口で表面の泥を食べる。ろ過摂食ではないと思う。赤い部分はとても食べる気はしないが,触手は食べられる。

図 31.干潟の環境と生物の調査(西表島の干立)。この辺りは海から 100~200m しか離れていないので,上げ潮に乗って海から大量の砂が入る。

図 32.干立の干潟で採集されたスナモグリ。スナモグリの種類に関してはやっと分かり始めた。近いうちに種類と系統分類に関する記事を出すだろう。

図 33.西表島の河口やマングローブの干潟に生息しているのはスナモグリが 2 種類,アナジャコが 3 種類ではなかろうか?アナジャコはすべてUpogebia や Gebia という属(genus)にまとめるのはまずい。分子系統学的研究の成果を含め,いくつかの新しい属を設けて,それらの中に仕分けするのが妥当ではないかと思う。また,分類学者の行うスケッチは客観的な研究手法とは言えず,検証不可能な結果をもたらすことに注意。